
1. Úvod
Jedním z dlouhodobých problémů ptačí fylogenetiky jsou příbuzenské vztahy pěvců. Tato skupina zahrnuje přes 60% terminálních taxonů ("druhů") žijících ptáků, její příbuzenství se zbytkem neoavianů (tj. všech ptáků krom paleognátů, hrabavých a vrubozobých) ale zůstávají otázkou. Morfologické analýzy dlouho navrhovaly jejich sesterskou pozici buďto vůči "srostloprstým" ("Coraciiformes"), nebo šplhavcům (Piciformes). Livezey & Zusi (2007) tyto tři taxony odkryli v polytomii – nedokázali určit, které dva formují skupinu s vyloučením toho třetího. Molekulární i morfologické rozbory se časem shodly na tom, že šplhavci ve skutečnosti patří dovnitř "srostloprstých". Společně tvoří klad, který Mayr (2011) nedávno nazval Picocoraciae. Fain & Houde (2004) odhalili na základě své kontroverzní fylogenetické studie 7. intronu β-fibrinogenu tento klad jako sesterskou skupinu pěvců. Jiné geny ale naznačovaly jiná příbuzenství: Sorenson et al. (2003) např. analyzovali 9. intron jaderného genu pro fosfoenolpyruvátovou karboxykinázu (PEPCK) a odhalili kuriózní sesterský vztah mezi pěvci a papoušky. O rok později jej ale potvrdila některá dílčí data předložená Cracraftem et al. (2004), konkrétně pak jaderný onkogen c-myc; a první fylogenomická analýza ptáků v pravém smyslu slova, kombinující údaje z pěti jaderných genů (Ericson et al. 2006), jej podpořila rovněž. Navrhla navíc, že celý spekulativní klad (papoušci + pěvci) má neméně neočekávané příbuzné, a to sokolovité (Falconidae) a seriemy.
2. Retropozony a proč jim věřit
Jedním z dlouhodobých problémů ptačí fylogenetiky jsou příbuzenské vztahy pěvců. Tato skupina zahrnuje přes 60% terminálních taxonů ("druhů") žijících ptáků, její příbuzenství se zbytkem neoavianů (tj. všech ptáků krom paleognátů, hrabavých a vrubozobých) ale zůstávají otázkou. Morfologické analýzy dlouho navrhovaly jejich sesterskou pozici buďto vůči "srostloprstým" ("Coraciiformes"), nebo šplhavcům (Piciformes). Livezey & Zusi (2007) tyto tři taxony odkryli v polytomii – nedokázali určit, které dva formují skupinu s vyloučením toho třetího. Molekulární i morfologické rozbory se časem shodly na tom, že šplhavci ve skutečnosti patří dovnitř "srostloprstých". Společně tvoří klad, který Mayr (2011) nedávno nazval Picocoraciae. Fain & Houde (2004) odhalili na základě své kontroverzní fylogenetické studie 7. intronu β-fibrinogenu tento klad jako sesterskou skupinu pěvců. Jiné geny ale naznačovaly jiná příbuzenství: Sorenson et al. (2003) např. analyzovali 9. intron jaderného genu pro fosfoenolpyruvátovou karboxykinázu (PEPCK) a odhalili kuriózní sesterský vztah mezi pěvci a papoušky. O rok později jej ale potvrdila některá dílčí data předložená Cracraftem et al. (2004), konkrétně pak jaderný onkogen c-myc; a první fylogenomická analýza ptáků v pravém smyslu slova, kombinující údaje z pěti jaderných genů (Ericson et al. 2006), jej podpořila rovněž. Navrhla navíc, že celý spekulativní klad (papoušci + pěvci) má neméně neočekávané příbuzné, a to sokolovité (Falconidae) a seriemy.
2. Retropozony a proč jim věřit
Výše uvedený text může naznačovat rostoucí shodu, a skutečně tomu tak v jistém ohledu je. Zároveň se ale objevovalo množství analýz, které spojitost mezi papoušky a pěvci nepotvrdily: především rozbory mitochondriální DNA mají tendence umisťovat druhou z těchto skupin na samou bázi kladu Neoaves (Slack et al. 2007); jiné analýzy tohoto typu dat ale zase podpořily příbuzenství se šplhavci (Gibb et al. 2007) či kukačkami (Pratt et al. 2009). Není proto divu, že systematiky začly lákat další zdroje znaků. Mezi nimi zaujímají prominentní místo retropozony: "skákající geny", repetitivní úseky DNA, které se začleňují do (téměř) náhodných míst genomu poté, co byly reverzně transkriptovány z nějaké molekuly RNA. Tyto úseky se dědí stejně jako kterýkoli jiný gen, vznik nového retropozonu je ovšem jedinečnou, neopakovatelnou událostí. Retropozony jsou poměrně dlouhé (v řádu stovek bází) a jejich inzerce je doprovázena tím, že se cílové místo duplikuje tak, aby vložený element původní sekvence lemovala z obou stran. Homoplazie by vyžadovala buď to, aby došlo k dokonale přesné deleci (zmizení) dříve vloženého retropozonu spolu s jednou kopií cílového místa, nebo naopak k inzerci stejného či extrémně podobného retropozonu na stejné místo genomu (přičemž možných míst inzerce je v ptačím genomu asi 1,2 miliardy). Pravděpodobnost obou variant je tak malá, že lze inzerci retropozonů řadit do kategorie tzv. vzácných genomových změn (RGC, Rare Genomic Changes), které jsou pro fylogenetika cosi jako splněný sen: dlouho přetrvávající znaky prakticky nevykazující homoplazii, ideální pro rozřešení dlouhotrvajících systematických kontroverzí.
Je nutné si uvědomit, že rozbor retropozonových inzercí je zcela nezávislý na dosavadních pokusech fylogeneticky analyzovat genetická data, ať už šlo o DNA-DNA hybridizaci nebo výše zmiňované sekvenční analýzy. To znamená, že analýzou retropozonů můžeme dosud navržené hypotézy testovat. Suh et al. (2011) považovali za příliš lákavé nechat si takovou možnost ujít. Vzali tedy genetický materiál kuřete (Gallus gallus) a zebřičky pestré (Taeniopygia guttata) – jediných dvou ptáků, jejichž genom byl kompletně přečten (osekvenován) –, přidali k němu známé sekvence pštrosa emu (Dromaius novaehollandiae) a nechali počítač najít v nich retropozony. Celkem jich rozeznali přes 200 000. Následně se zaměřili na dvě jejich nejrozsáhlejší kategorie, zahrnující přes 97% z celkového množství: kuřecí repetice 1 (CR1), což je rodina dlouhých vmezeřených elementů (LINEs, Long Interspersed Nuclear Elements), a dlouhé koncové repetice (LTRs, Long Terminal Repeats) endogenních retrovirů. Z nich potom vybrali ty retropozony, které jsou součástí zachovaných (tedy pomalu se vyvíjejících a snadno rozeznatelných) intronů a mezigenových oblastí o velikosti menší než 1500 párů bází.
Tyto podmínky splnilo 131 lokusů s dlouhými vmezeřenými elementy a 75 lokusů s dlouhými koncovými repeticemi. Většina z nich byla k nalezení u zebřičky, ale ne u kuřete či emu, čímž se prokázalo, že právě tyto elementy jsou relevantní k rozřešení příbuzenských vztahů mezi neoaviany. Pomocí amplifikace polymerázovou řetězovou reakcí (PCR) a sekvenování se pokusili tyto sekvence získat i od jiných ptáků: od papouška kea (Nestor), sokola (Falco), žluny (Picus), káněte (Buteo), čápa (Ciconia) a holuba (Columba). To se ukázalo být možné pro 51 z celkových 206 původních kandidátů. Suh et al. (2011) sekvence těchto retropozonů zarovnali a sestavili z nich datovou matrici. Každý taxon může vykazovat buď primitivní, ancestrální stav (v ortologním* lokusu konkrétní retropozon chybí), kódovaný jako "–"; nebo naopak stav odvozený (došlo k inzerci příslušného retropozonu) kódovaný jako "+". Třetí možností je, že v genomu ptáka cílové místo pro konkrétní retropozon kvůli deleci zcela chybělo. Pak jej Suh et al. (2011) kódovali jako "d" (chybějící data). Následně použili PAUP k tomu, aby z matrice zrekonstruoval kladogram. Vzali přitom v úvahu návrh Waddella et al. (2001), kteří navrhli zkolabovat každou větev, která není podložena nejméně třemi navzájem nekonfliktními retropozony. (Návrh pramení z toho, že ač je konvergence u retropozonů extrémně nepravděpodobná, vyloučená není, a jeden nebo dva retropozony tedy nejsou statisticky signifikantní podporou). Výsledky analýzy vypadají následovně:
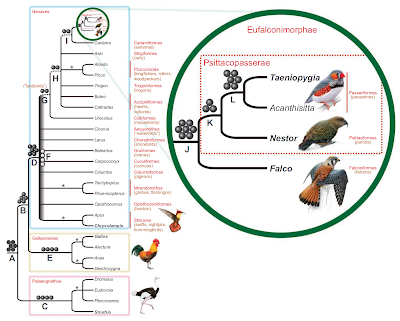
Je nutné si uvědomit, že rozbor retropozonových inzercí je zcela nezávislý na dosavadních pokusech fylogeneticky analyzovat genetická data, ať už šlo o DNA-DNA hybridizaci nebo výše zmiňované sekvenční analýzy. To znamená, že analýzou retropozonů můžeme dosud navržené hypotézy testovat. Suh et al. (2011) považovali za příliš lákavé nechat si takovou možnost ujít. Vzali tedy genetický materiál kuřete (Gallus gallus) a zebřičky pestré (Taeniopygia guttata) – jediných dvou ptáků, jejichž genom byl kompletně přečten (osekvenován) –, přidali k němu známé sekvence pštrosa emu (Dromaius novaehollandiae) a nechali počítač najít v nich retropozony. Celkem jich rozeznali přes 200 000. Následně se zaměřili na dvě jejich nejrozsáhlejší kategorie, zahrnující přes 97% z celkového množství: kuřecí repetice 1 (CR1), což je rodina dlouhých vmezeřených elementů (LINEs, Long Interspersed Nuclear Elements), a dlouhé koncové repetice (LTRs, Long Terminal Repeats) endogenních retrovirů. Z nich potom vybrali ty retropozony, které jsou součástí zachovaných (tedy pomalu se vyvíjejících a snadno rozeznatelných) intronů a mezigenových oblastí o velikosti menší než 1500 párů bází.
Tyto podmínky splnilo 131 lokusů s dlouhými vmezeřenými elementy a 75 lokusů s dlouhými koncovými repeticemi. Většina z nich byla k nalezení u zebřičky, ale ne u kuřete či emu, čímž se prokázalo, že právě tyto elementy jsou relevantní k rozřešení příbuzenských vztahů mezi neoaviany. Pomocí amplifikace polymerázovou řetězovou reakcí (PCR) a sekvenování se pokusili tyto sekvence získat i od jiných ptáků: od papouška kea (Nestor), sokola (Falco), žluny (Picus), káněte (Buteo), čápa (Ciconia) a holuba (Columba). To se ukázalo být možné pro 51 z celkových 206 původních kandidátů. Suh et al. (2011) sekvence těchto retropozonů zarovnali a sestavili z nich datovou matrici. Každý taxon může vykazovat buď primitivní, ancestrální stav (v ortologním* lokusu konkrétní retropozon chybí), kódovaný jako "–"; nebo naopak stav odvozený (došlo k inzerci příslušného retropozonu) kódovaný jako "+". Třetí možností je, že v genomu ptáka cílové místo pro konkrétní retropozon kvůli deleci zcela chybělo. Pak jej Suh et al. (2011) kódovali jako "d" (chybějící data). Následně použili PAUP k tomu, aby z matrice zrekonstruoval kladogram. Vzali přitom v úvahu návrh Waddella et al. (2001), kteří navrhli zkolabovat každou větev, která není podložena nejméně třemi navzájem nekonfliktními retropozony. (Návrh pramení z toho, že ač je konvergence u retropozonů extrémně nepravděpodobná, vyloučená není, a jeden nebo dva retropozony tedy nejsou statisticky signifikantní podporou). Výsledky analýzy vypadají následovně:

Striktní konsenzus 577 nejúspornějších stromků, vzešlých z kladistické analýzy retropozonových inzercí. Černá kolečka (pouze nad větví A) reprezentují inzerce (vložení) retropozonů, které jsou jedinečné pro ptáky jako celek; tmavošedé kuličky (např. nad větví B) symbolizují inzerce se shodným výskytem, které dávají shodný fylogenetický signál. Světle šedé kuličky (větev F) zastupují retropozony, které poskytují signály rozporné mezi sebou a s retropozonovými markery na čárkovaných větvích. Podle autorů jde o následek tzv. ILS (Incomplete Lineage Sorting) efektu (viz dále v textu), o němž je známo, že může způsobit nekompatibilitu mezi genovými genealogiemi a celkovou fylogenezí. Některé z tradičních a dobře podložených kladů se překvapivě ukázaly být bez retropozonové podpory: sem patří nejen stále trochu kontroverzní mirandorniti (plameňáci + potápky) a nepojmenovaný klad-tvořený-všemi-paleognáty-kromě-pštrosa, ale i hrabaví, vrubozobí, svišťouni nebo Picocoraciae (šplhavci + parafyletičtí "srostloprstí"). Suh et al. (2011) takové skupiny označili na výsledném kladogramu hvězdičkou. Na stromku se mi také líbí to, jak pohotově do něj autoři zařadili teprve nedávno navržená, ale vysoce užitečná jména; především pak ta z Mayrovy letošní práce – Aequornithes a Picocoraciae (Mayr 2011). (Zdroj: Suh et al. 2011: Figure 1)
O tom, že retropozony jsou spolehlivými indikátory fylogeneze, vypovídá už fakt, že jejich analýza potvrdila většinu tradičních seskupení už dříve potvrzených morfologickými i sekvenčními daty; mezi nimi pěvce (Passeriformes), drůbež (Galloanserae), monofylii neoavianů a dělení moderních ptáků na Palaeognathae a Neognathae. Pak už můžeme přistoupit k tomu, co nám retropozony říkají o problémech, které morfologie a ani sekvenční rozbory dosud nedokázaly uspokojivě vyřešit – o bazálních divergencích skupiny Neoaves.
3. Neoaves – měkká, nebo tvrdá polytomie?
Špatnou zprávou je, že genealogie čtyř retropozonových markerů s potenciálem rozřešit neoavianní vývojovou radiaci se ukázala být vzájemně nekonzistentní. Již bylo vysvětleno, že konvergence je v případě retropozonů na hranici nemožného: vložení stejného typu retropozonu, stejně zkráceného (truncated) a stejně orientovaného, na přesně stejné místo v genomu je vyloučena zákony pravděpodobnosti, a dokonale přesné "vystříhnutí" takového elementu by (bez ohledu na statistickou nepravděpodobnost) muselo být rozpoznáno. Suh s kolektivem proto nesrovnalosti mezi jednotlivými elementy připisují jevu zvanému "incomplete lineage sorting". Po oddělení dvou populací genetický drift v každé z nich stochasticky (náhodně) zafixuje různé alely, k tomu ovšem potřebuje nějaký čas: konkrétně 2Ne generací (kde Ne znamená efektivní velikost populace). Do té doby si obě linie, které se zvolna stávají novými taxony, udržují polymorfizmus zděděný od ancestrální populace, což znamená, že jeden lokus obsazují různé alely. Problém nastává v okamžiku, pokud se jedna z těchto linií znovu rozštěpí dřív, než genetický drift fixuje v polymorfním lokusu jednu alelu. Pak se polymorfizmus přenese do nové dvojice linií, a drift v nich potom může fixovat alely takovým způsobem, který odporuje celkové fylogenezi. Protože jeden obrázek často řekne víc, než 106 slov, kterými jsem právě zkusil incomplete lineage sorting vysvětlit, ukradnu jeden od Aviseho a Robinsona (2008), kteří pro podobné těžkosti zavedli pojem hemiplazie:

Polymorfizmus z doby před oddělením větví (A + B) a C přetrval až do doby, dokud se (A + B) sama nerozdělila na A a B, a teprve potom se fixoval takovým způsobem, který odporuje celkovému uspořádání ("species tree"). Je nutné zdůraznit, jak to činí i Avise & Robinson (2008), že genový strom není špatně. Z hlediska "species tree" nám to tak může připadat, ale v genovém stromu je (B + C) monofyletickou skupinou a fixace alely b validní homologií. Genealogie genů se v tomto případě odlišuje od genealogie taxonů, jejichž zástupce geny "obývají", což ale neznamená, že by jedna z nich musela být nutně špatně. Pěkně celou situaci vysvětluje např. Dawkins (2004): každý gen má vlastní genealogii, která se může lišit jak od genealogií jiných genů, tak i od fylogeneze organizmů, které autorův pohled zredukoval na pouhé vehikly pro velké množství genů. "Species tree" je prostě ta topologie, na které se shodne většina genů v demokratickém hlasování. (Zdroj: Avise & Robinson 2008: Figure 2)
Hemiplazie je důležitá právě proto, že zasahuje i vzácné genomové změny. I kdyby byly všechny jevy z této kategorie skutečně dokonale prosté jakékoli homoplazie, díky hemiplazii se fylogeneze z nich odvozená stále může odlišovat od té, kterou hledáme (tj. od "species tree"). Stačí, aby měla báze kladu, jehož topologii se snažíme odhalit, větve o velmi krátké délce. Ve fylogenetice se pro taková místa ujalo neformální označení "bush" (křoví) jako variace na klasickou představu o fylogenezi, kterou ztělesňuje strom ("tree"); a jsou důvody se domnívat, že nejsou nijak vzácná (Rokas & Carroll 2006).
Jedním z takových míst by mohla být právě báze kladu Neoaves. Poe & Chubb (2004) přišli se značně pesimistickým modelem "explozivní radiace", podle nějž je tato oblast stromu života tzv. "tvrdou polytomií", která – na rozdíl od polytomií měkkých, které jsou jen projevem nedostatku údajů spíše než fundamentálních metodologických překážek – nebude rozřešena ani sebevětším množstvím genetických dat. Mělo by jít o jedno z právě těch míst, o kterých psali Rokas a Carroll, kde ani stovky genů nedokážou poskytnout dostatečné rozlišení. Zdá se, že neshoda v retropozonových markerech, kterou zjistili Suh et al. (2011) a za kterou je zřejmě zodpovědná právě neschopnost genetického driftu zafixovat ve čtyřech problematických lokusech různé alely před tím, než se větev začala znovu štěpit, tento model pouze potvrzuje. Je ale situace skutečně tak bledá?
Chojnowski et al. (2008) byli optimističtější. Ukázali, že ačkoli exony (kódující místa genů) skutečně nemusejí na rozřešení stromů s velice krátkými meziuzly (= "internodes" nebo prostě "větve"; myslí se tím úsek mezi dvěma divergencemi) stačit, introny by to zvládnout mohly, neboť v genových stromech odvozených z intronů dvou paralogních genů pro těžké řetězce klatrinu našli větší shodu, než jakou je možné vysvětlit čistou náhodou. Nadšení ale v květnu letošního roku zchladili Braun et al. (2011), kteří se zaměřili na další ze vzácných genomových změn: mikroinverze, tedy převrácení posloupnosti několika desítek až několika desítek tisíc bází. Jednak ukázali, že mikroinverze zase tak vzácné nejsou a vykazují homoplazii, kterou nelze vysvětlit výše popsaným mechanizmem hemiplazie; jednak spočítali, že k rozřešení meziuzlů o délce 1 milionu let pomocí mikroinverzí by bylo třeba prohledat kolem 12 milionů bází, což je o tři řády více, než používají sekvenční analýzy typu Hackett et al. (2008). Existují přitom doklady, že meziuzly na samé bázi Neoaves jsou možná ještě kratší, a to navíc stále předpokládáme, že tato oblast představuje polytomii měkkou, o čemž Braun et al. (2011) sami vyjádřili nejistotu.
4. Psittacopasserae a Eufalconimorphae
Další potvrzení problematičnosti hlubokých příbuzenských vztahů v rámci Neoaves ale pochopitelně není to hlavní, co si ze studie Suha a kolektivu máme odnést. Jak autoři zmínili přímo ve své práci a jak ještě opětovně zdůraznil Alexander Suh v e-mailové konverzaci na DML (do které jsem měl tu čest se zapojit), hlavní je, že rozbor nového typu dat potvrdil opatrný konsenzus, který se začal rýsovat s nástupem fylogenomických studií v posledních letech. Jedním z kladů navržených sekvenčními analýzami, který má oporu také v retropozonech, jsou tzv. pozemní ptáci (o jejichž historii plánuji ještě něco napsat.) Vynořil se poprvé s velmi slušnou podporou ze studie Ericsona et al. (2006) a zahrnuje pěvce, papoušky, sokolovité, seriemy, denní dravce, sovy, myšáky, trogony, madagaskarského kurola (Leptosomus) a pikokoraciany (zoborožci, dudci, leskovci, datli, ledňáčci, vlhovití atd.). Morfologové zdánlivě podobná seskupení navrhovali již dříve, ale do moderního pojetí jim buď cosi chybělo (denní dravci, papoušci, seriemy), nebo naopak přebývalo – hlavně různé kukačky, svišťouni a "lelci" (srovnej např. "Dendrornithes" z Livezey & Zusi 2007 a "landbirds" z Hackett et al. 2008). Molekulární analýzy jsou přitom nekompromisní: Hackett et al. (2008) podložili "pozemní ptáky" s výše popsaným obsahem bootstrapovou podporu 98% a Suh et al. (2011) nyní jejich monofylii potvrzují znovu, ačkoli podle vlastních kritérií nedostatečně: našli jen 2 retropozonové inzerce místo 3 a navíc jim odporují některé ze čtyř výše zmíněných hemiplastických markerů u báze Neoaves (viz popisek k prvnímu obrázku).
Úplně nový a dosud nikde nenavržený je další klad, sestávající ze všech pozemních ptáků kromě myšáků (Coliidae). Podpora pro něj je stejná, jako v případě pozemních ptáků samotných: dvě inzerce, které jsou v rozporu s několika dalšími markery. Přesto jde o hypotézu, která si zaslouží pozornost a další testovaní. Pozici myšáků nepomohly stabilizovat ani velké morfologické či sekvenční analýzy posledních let (jen stručně: Chubb [2004] navrhl sesterský vztah s papoušky, Livezey & Zusi [2007] s trogony a Hackett et al. [2008] se sovami), díky čemuž si tato skupinka drobných afrických ptáků buduje reputaci fylogenetické hádanky srovnatelné i s proslulým hoacinem.
Situace se začíná lepšit s tím, jak postupujeme k méně rozsáhlým skupinám. Klad složený ze seriem, sokolů, papoušků a pěvců* sice také není podložený zvlášť dobře, ale v kombinaci s podporou, jakou pro něj přinesly velké sekvenční studie (posteriorní pravděpodobnost přes 95% v práci Ericsona a kolektivu), se jeho existence zdá být velmi pravděpodobná. Jeho vnitřní příbuzenství jsou již retropozonovými daty zdokumentována velmi dobře: klad zahrnující papoušky, pěvce a sokolovité je dokonce nejlépe podloženým seskupením v celé analýze. Podporuje jej 7 retropozonových inzercí (všechno dlouhé koncové repetice odvozené z endogenních retrovirů) a – na rozdíl od výše zmíněných příbuzenství – neodporuje jim ani jeden ze 196 zbývajících markerů; což je evidence, které nemohly konkurovat ani tak tradiční klady, jako jsou neognáti, paleognáti, drůbež nebo pěvci. Suh et al. (2011) se proto cítili dostatečně oprávněni k tomu, aby jej pojmenovali. Zvolili "Eufalconimorphae", což je označení, které na DML vyvolalo rozsáhlou diskuzi. Většina komentářů jej přitom hodnotila jako příšerné. Osobně jsem se snažil argumentovat, že název jistou logiku má – předpona "eu-", čili "praví", poukazuje na to, že sem nepatří žádní z tradičních falkoniformů, jako jsou jestřábovití, orlovci nebo hadilov – a Alexander Suh později v odpovědi na můj e-mail tuto spekulaci potvrdil.
*Tento klad nemá žádné jméno, a ani Suh et al. (2011) pro něj žádné vzhledem k nízké podpoře ze strany retropozonů nenavrhli. Boyd (2011) používá "Passerimorphae", což je, jak jsem pochopil, neustále recyklovaný výraz pro pěvce a dva až tři "řády" zrovna považované za jejich nejbližší příbuzné. Přišli s ním Sibley et al. (1988), podle nichž byly těmito "řády" holubi, "krátkokřídlí" a mimořádně inkluzivní verze "brodivých"; Livezey & Zusi (2007) jej použili pro pěvce, "srostloprsté" a šplhavce.
V rámci eufalkonimorfů jsou pak zřejmě sokolovití (Falconidae) sesterskou skupinou kladu složeného z pěvců (Passeriformes) a papoušků (Psittaciformes). Tento klad je podložen třemi retropozony, kterým opět žádný ze zbývajících neodporuje. Jelikož sesterský vztah mezi oběma skupinami podporují také sekvence hned tří jaderných genů (Mayr 2011), rozhodli se Suh et al. (2011) pojmenovat i tento klad: Psittacopasserae. Upozorňují nicméně na nutnost dalších sekvenčních analýz, protože ačkoli nenašli žádné známky po ILS, nemohou zcela vyloučit hemiplazii v případě retropozonů spojujících papoušky a pěvce.
Autoři se dále zaměřili na evoluční historii ptačích retropozonů. Vzali si na pomoc analýzu Pereiry a Bakera (2006), která načasování zásadních ptačích divergencí neodvodila pomocí běžných metod, jako jsou molekulární hodiny, ale bayesovskou pravděpodobnostní analýzou nad kladogramem zrekonstruovaným z mitochondriální DNA. Poté se podívali na takové retropozony, které jsou usazeny uvnitř jiných retropozonů – v genomu zebřičky pestré jich našli 995. Za předpokladu, že mladší (aktivní) retropozony se častěji vkládají do starších (neaktivních), spíš než aby tomu bylo naopak, může analýza zjistit, jaké typy těchto elementů byly aktivní v jakou dobu. Povedlo se např. identifikovat retropozony aktivní v období, než došlo k oddělení kuřecí linie (Galloanserae) od linie zebřičky (Neoaves), a také elementy, které byly aktivní v období neoavianní vývojové radiace. Zvláštní je, že jde hlavně o dlouhé koncové repetice s virovým původem, což naznačuje, že se toto období explozivního vzniku nových taxonů krylo s obdobím zvýšené aktivity endogenních retrovirů. Warren et al. (2010), kteří vloni oznámili přečtení kompletního genomu zebřičky pestré, v něm skutečně identifikovali třikrát víc dlouhých koncových repeticí, než je známo z genomu kuřete. Suh et al. (2011) zdůrazňují, že jejich datace retropozonů může významně usnadnit práci autorům budoucích analýz – dovolí jim zaměřit se přesně na ty elementy, které jsou relevantní pro právě zkoumaný úsek evoluční historie ptáků.
5. Kolikrát se ptáci naučili učit se zpívat?
Autoři také diskutují o jedné z nejdůležitějších implikací blízkého příbuzenství papoušků a pěvců. Obě skupiny jsou totiž charakteristické svou vokalizací (hlasovými projevy): papoušci se proslavili svou imitací lidské řeči a pěvci dokonce po této schopnosti dostali své jméno – ačkoli vlastně nepřesné. Zpěv, který je naučený a nikoli výhradně vrozený, je totiž zaznamenán jen u jedné (ačkoli zdaleka největší) podskupiny pěvců, zvané Oscines (česky "zpěvní"). O sesterské skupině zpěvných, křikavých (Suboscines), a sesterské skupině celého kladu (Oscines + Suboscines), novozélandských pokřovníkovitých (Acathisittidae), kteří jsou obecně prozkoumaní velice málo, toho totiž z hlediska vokalizace příliš nevíme. Ví se, že někteří křikaví Nového světa (zástupci skupiny Tyrannides), např. tyranovití, skutečně nejsou vokálního učení schopni; zatímco jiní zástupci téhož kladu, např. kotingovití (Cotingidae) a pipulkovití (Pipridae) ano. Suhovi a spol. proto oprávněně připadá úspornější hypotéza, že hlasové učení vzniklo jen jednou u společného předka kladu Psittacopasserae a následně bylo několikrát ztraceno, než aby se u blízce příbuzných taxonů vyvíjelo několikrát nezávisle na sobě. To by znamenalo, že mozkové oblasti a vzorce genové exprese, zjištěné odděleně u papoušků a (zpěvných) pěvců – hlavně pak tzv. přední hlasová dráha v předním mozku – jsou rovněž homologické, a že jejich vznik musí být posunut do minulosti až o 30 milionů let oproti předcházejícím odhadům, které předpokládaly jejich konvergentní vývoj. Suh et al. (2011) uvádějí, že důsledky nově podloženého příbuzenství by mohly být ještě závažnější a změnit náš celkový pohled na evoluci hlasového učení.

Zdroje:Jedním z takových míst by mohla být právě báze kladu Neoaves. Poe & Chubb (2004) přišli se značně pesimistickým modelem "explozivní radiace", podle nějž je tato oblast stromu života tzv. "tvrdou polytomií", která – na rozdíl od polytomií měkkých, které jsou jen projevem nedostatku údajů spíše než fundamentálních metodologických překážek – nebude rozřešena ani sebevětším množstvím genetických dat. Mělo by jít o jedno z právě těch míst, o kterých psali Rokas a Carroll, kde ani stovky genů nedokážou poskytnout dostatečné rozlišení. Zdá se, že neshoda v retropozonových markerech, kterou zjistili Suh et al. (2011) a za kterou je zřejmě zodpovědná právě neschopnost genetického driftu zafixovat ve čtyřech problematických lokusech různé alely před tím, než se větev začala znovu štěpit, tento model pouze potvrzuje. Je ale situace skutečně tak bledá?
Chojnowski et al. (2008) byli optimističtější. Ukázali, že ačkoli exony (kódující místa genů) skutečně nemusejí na rozřešení stromů s velice krátkými meziuzly (= "internodes" nebo prostě "větve"; myslí se tím úsek mezi dvěma divergencemi) stačit, introny by to zvládnout mohly, neboť v genových stromech odvozených z intronů dvou paralogních genů pro těžké řetězce klatrinu našli větší shodu, než jakou je možné vysvětlit čistou náhodou. Nadšení ale v květnu letošního roku zchladili Braun et al. (2011), kteří se zaměřili na další ze vzácných genomových změn: mikroinverze, tedy převrácení posloupnosti několika desítek až několika desítek tisíc bází. Jednak ukázali, že mikroinverze zase tak vzácné nejsou a vykazují homoplazii, kterou nelze vysvětlit výše popsaným mechanizmem hemiplazie; jednak spočítali, že k rozřešení meziuzlů o délce 1 milionu let pomocí mikroinverzí by bylo třeba prohledat kolem 12 milionů bází, což je o tři řády více, než používají sekvenční analýzy typu Hackett et al. (2008). Existují přitom doklady, že meziuzly na samé bázi Neoaves jsou možná ještě kratší, a to navíc stále předpokládáme, že tato oblast představuje polytomii měkkou, o čemž Braun et al. (2011) sami vyjádřili nejistotu.
4. Psittacopasserae a Eufalconimorphae
Další potvrzení problematičnosti hlubokých příbuzenských vztahů v rámci Neoaves ale pochopitelně není to hlavní, co si ze studie Suha a kolektivu máme odnést. Jak autoři zmínili přímo ve své práci a jak ještě opětovně zdůraznil Alexander Suh v e-mailové konverzaci na DML (do které jsem měl tu čest se zapojit), hlavní je, že rozbor nového typu dat potvrdil opatrný konsenzus, který se začal rýsovat s nástupem fylogenomických studií v posledních letech. Jedním z kladů navržených sekvenčními analýzami, který má oporu také v retropozonech, jsou tzv. pozemní ptáci (o jejichž historii plánuji ještě něco napsat.) Vynořil se poprvé s velmi slušnou podporou ze studie Ericsona et al. (2006) a zahrnuje pěvce, papoušky, sokolovité, seriemy, denní dravce, sovy, myšáky, trogony, madagaskarského kurola (Leptosomus) a pikokoraciany (zoborožci, dudci, leskovci, datli, ledňáčci, vlhovití atd.). Morfologové zdánlivě podobná seskupení navrhovali již dříve, ale do moderního pojetí jim buď cosi chybělo (denní dravci, papoušci, seriemy), nebo naopak přebývalo – hlavně různé kukačky, svišťouni a "lelci" (srovnej např. "Dendrornithes" z Livezey & Zusi 2007 a "landbirds" z Hackett et al. 2008). Molekulární analýzy jsou přitom nekompromisní: Hackett et al. (2008) podložili "pozemní ptáky" s výše popsaným obsahem bootstrapovou podporu 98% a Suh et al. (2011) nyní jejich monofylii potvrzují znovu, ačkoli podle vlastních kritérií nedostatečně: našli jen 2 retropozonové inzerce místo 3 a navíc jim odporují některé ze čtyř výše zmíněných hemiplastických markerů u báze Neoaves (viz popisek k prvnímu obrázku).
Úplně nový a dosud nikde nenavržený je další klad, sestávající ze všech pozemních ptáků kromě myšáků (Coliidae). Podpora pro něj je stejná, jako v případě pozemních ptáků samotných: dvě inzerce, které jsou v rozporu s několika dalšími markery. Přesto jde o hypotézu, která si zaslouží pozornost a další testovaní. Pozici myšáků nepomohly stabilizovat ani velké morfologické či sekvenční analýzy posledních let (jen stručně: Chubb [2004] navrhl sesterský vztah s papoušky, Livezey & Zusi [2007] s trogony a Hackett et al. [2008] se sovami), díky čemuž si tato skupinka drobných afrických ptáků buduje reputaci fylogenetické hádanky srovnatelné i s proslulým hoacinem.
Situace se začíná lepšit s tím, jak postupujeme k méně rozsáhlým skupinám. Klad složený ze seriem, sokolů, papoušků a pěvců* sice také není podložený zvlášť dobře, ale v kombinaci s podporou, jakou pro něj přinesly velké sekvenční studie (posteriorní pravděpodobnost přes 95% v práci Ericsona a kolektivu), se jeho existence zdá být velmi pravděpodobná. Jeho vnitřní příbuzenství jsou již retropozonovými daty zdokumentována velmi dobře: klad zahrnující papoušky, pěvce a sokolovité je dokonce nejlépe podloženým seskupením v celé analýze. Podporuje jej 7 retropozonových inzercí (všechno dlouhé koncové repetice odvozené z endogenních retrovirů) a – na rozdíl od výše zmíněných příbuzenství – neodporuje jim ani jeden ze 196 zbývajících markerů; což je evidence, které nemohly konkurovat ani tak tradiční klady, jako jsou neognáti, paleognáti, drůbež nebo pěvci. Suh et al. (2011) se proto cítili dostatečně oprávněni k tomu, aby jej pojmenovali. Zvolili "Eufalconimorphae", což je označení, které na DML vyvolalo rozsáhlou diskuzi. Většina komentářů jej přitom hodnotila jako příšerné. Osobně jsem se snažil argumentovat, že název jistou logiku má – předpona "eu-", čili "praví", poukazuje na to, že sem nepatří žádní z tradičních falkoniformů, jako jsou jestřábovití, orlovci nebo hadilov – a Alexander Suh později v odpovědi na můj e-mail tuto spekulaci potvrdil.
*Tento klad nemá žádné jméno, a ani Suh et al. (2011) pro něj žádné vzhledem k nízké podpoře ze strany retropozonů nenavrhli. Boyd (2011) používá "Passerimorphae", což je, jak jsem pochopil, neustále recyklovaný výraz pro pěvce a dva až tři "řády" zrovna považované za jejich nejbližší příbuzné. Přišli s ním Sibley et al. (1988), podle nichž byly těmito "řády" holubi, "krátkokřídlí" a mimořádně inkluzivní verze "brodivých"; Livezey & Zusi (2007) jej použili pro pěvce, "srostloprsté" a šplhavce.
V rámci eufalkonimorfů jsou pak zřejmě sokolovití (Falconidae) sesterskou skupinou kladu složeného z pěvců (Passeriformes) a papoušků (Psittaciformes). Tento klad je podložen třemi retropozony, kterým opět žádný ze zbývajících neodporuje. Jelikož sesterský vztah mezi oběma skupinami podporují také sekvence hned tří jaderných genů (Mayr 2011), rozhodli se Suh et al. (2011) pojmenovat i tento klad: Psittacopasserae. Upozorňují nicméně na nutnost dalších sekvenčních analýz, protože ačkoli nenašli žádné známky po ILS, nemohou zcela vyloučit hemiplazii v případě retropozonů spojujících papoušky a pěvce.
Autoři se dále zaměřili na evoluční historii ptačích retropozonů. Vzali si na pomoc analýzu Pereiry a Bakera (2006), která načasování zásadních ptačích divergencí neodvodila pomocí běžných metod, jako jsou molekulární hodiny, ale bayesovskou pravděpodobnostní analýzou nad kladogramem zrekonstruovaným z mitochondriální DNA. Poté se podívali na takové retropozony, které jsou usazeny uvnitř jiných retropozonů – v genomu zebřičky pestré jich našli 995. Za předpokladu, že mladší (aktivní) retropozony se častěji vkládají do starších (neaktivních), spíš než aby tomu bylo naopak, může analýza zjistit, jaké typy těchto elementů byly aktivní v jakou dobu. Povedlo se např. identifikovat retropozony aktivní v období, než došlo k oddělení kuřecí linie (Galloanserae) od linie zebřičky (Neoaves), a také elementy, které byly aktivní v období neoavianní vývojové radiace. Zvláštní je, že jde hlavně o dlouhé koncové repetice s virovým původem, což naznačuje, že se toto období explozivního vzniku nových taxonů krylo s obdobím zvýšené aktivity endogenních retrovirů. Warren et al. (2010), kteří vloni oznámili přečtení kompletního genomu zebřičky pestré, v něm skutečně identifikovali třikrát víc dlouhých koncových repeticí, než je známo z genomu kuřete. Suh et al. (2011) zdůrazňují, že jejich datace retropozonů může významně usnadnit práci autorům budoucích analýz – dovolí jim zaměřit se přesně na ty elementy, které jsou relevantní pro právě zkoumaný úsek evoluční historie ptáků.
5. Kolikrát se ptáci naučili učit se zpívat?
Autoři také diskutují o jedné z nejdůležitějších implikací blízkého příbuzenství papoušků a pěvců. Obě skupiny jsou totiž charakteristické svou vokalizací (hlasovými projevy): papoušci se proslavili svou imitací lidské řeči a pěvci dokonce po této schopnosti dostali své jméno – ačkoli vlastně nepřesné. Zpěv, který je naučený a nikoli výhradně vrozený, je totiž zaznamenán jen u jedné (ačkoli zdaleka největší) podskupiny pěvců, zvané Oscines (česky "zpěvní"). O sesterské skupině zpěvných, křikavých (Suboscines), a sesterské skupině celého kladu (Oscines + Suboscines), novozélandských pokřovníkovitých (Acathisittidae), kteří jsou obecně prozkoumaní velice málo, toho totiž z hlediska vokalizace příliš nevíme. Ví se, že někteří křikaví Nového světa (zástupci skupiny Tyrannides), např. tyranovití, skutečně nejsou vokálního učení schopni; zatímco jiní zástupci téhož kladu, např. kotingovití (Cotingidae) a pipulkovití (Pipridae) ano. Suhovi a spol. proto oprávněně připadá úspornější hypotéza, že hlasové učení vzniklo jen jednou u společného předka kladu Psittacopasserae a následně bylo několikrát ztraceno, než aby se u blízce příbuzných taxonů vyvíjelo několikrát nezávisle na sobě. To by znamenalo, že mozkové oblasti a vzorce genové exprese, zjištěné odděleně u papoušků a (zpěvných) pěvců – hlavně pak tzv. přední hlasová dráha v předním mozku – jsou rovněž homologické, a že jejich vznik musí být posunut do minulosti až o 30 milionů let oproti předcházejícím odhadům, které předpokládaly jejich konvergentní vývoj. Suh et al. (2011) uvádějí, že důsledky nově podloženého příbuzenství by mohly být ještě závažnější a změnit náš celkový pohled na evoluci hlasového učení.

Obrázek zachycuje příbuzenské vztahy a stavbu mozku těch ptáků, kteří jsou schopni hlasového učení: kolibříků (hummingbirds), papoušků (parrots) a zpěvných pěvců (oscine passerines), spolu s jejich nejbližšími příbuznými, kteří tuto schopnost postrádají – rorýsovitými (swifts) a sokolovitými (falcons). Schémata mozku jsou převzata z Jarvise et al. (2000) a zachycují přední (červeně) a zadní (zeleně) hlasovou dráhu spolu se zadní drahou sluchovou, centrem sluchových reflexů (modře); jejich umístění je přitom odvozeno z míst, kde dochází k exprimaci genu ZENK. U rorýsovitých a sokolovitých potřebný výzkum ještě nebyl proveden, proto u nich schémata mozku chybějí. Černý čtverec značí odvozený znak, bílý potom znak primitivní, zděděný (pleziomorfický). Do budoucna je nutné prozkoumat neuroanatomii křikavých a pokřovníkovitých, aby se zjistilo, zda je zadní hlasová dráha papoušků a zpěvných zděděná od společného předka, nebo konvergentně vyvinutá. Proužek měřítka u mozkových schémat odpovídá 2 mm. (Zdroj: Suh et al. 2011: Figure 4)
- http://dml.cmnh.org/2011Aug/msg00401.html
- http://dml.cmnh.org/2011Sep/msg00004.html
- Avise JC, Robinson TJ 2008 Hemiplasy: a new term in the lexicon of phylogenetics. Syst Biol 57(3): 503–7
- Boyd JH III 2011 Taxonomy in Flux Checklist 2.52: Tree View. Accessed September 13, 2011 at http://jboyd.net/Taxo/tree_aves.html
- Braun EL, Kimball RT, Han K-L, Iuhasz-Velez NR, Bonilla AJ, Chojnowski JL, Smith JV, Bowie RCK, Braun MJ, Hackett SJ, Harshman J, Huddleston CJ, Marks BD, Miglia KJ, Moore WS, Reddy S, Sheldon FH, Witt CC, Yuri T 2011 Homoplastic microinversions and the avian tree of life. BMC Evol Biol 11: 141
- Chojnowski JL, Kimball RT, Braun EL 2008 Introns outperform exons in analyses of basal avian phylogeny using clathrin heavy chain genes. Gene 410: 89–96
- Chubb AL 2004 New nuclear evidence for the oldest divergence among neognath birds: the phylogenetic utility of ZENK (i). Mol Phylog Evol 30(1): 140–51
- Cracraft J, Barker FK, Braun M, Harshman J, Dyke GJ, Feinstein J, Stanley S, Cibois A, Schikler P, Beresford P, García-Moreno J, Sorenson MD, Yuri T, Mindell DP 2004 Phylogenetic relationships among modern birds (Neornithes): toward an avian tree of life. 468–89 in Cracraft J, Donoghue M, eds. Assembling the Tree of Life. New York: Oxford Univ Press
- Ericson PGP, Anderson CL, Britton T, Elżanowski A, Johansson US, Källersjö M, Ohlson JI, Parsons TJ, Zuccon D, Mayr G 2006 Diversification of Neoaves: Integration of molecular sequence data and fossils. Biol Lett 2(4): 543–7
- Fain MG, Houde P 2004 Parallel radiations in the primary clades of birds. Evolution 58(11): 2558–73
- Gibb GC, Kardailsky O, Kimball RT, Braun E, Penny D 2007 Mitochondrial genomes and avian phylogeny: complex characters and resolvability without explosive radiations. Mol Biol Evol 24: 269–80
- Hackett SJ, Kimball RT, Reddy S, Bowie RC, Braun EL, Braun MJ, Chojnowski JL, Cox WA, Han K, Harshman J, Huddleston CJ, Marks BD, Miglia KJ, Moore WS, Sheldon FH, Steadman DW, Witt CC, Yuri T 2008 A phylogenomic study of birds reveals their evolutionary history. Science 320(5884): 1763–8
- Jarvis ED, Ribeiro S, da Silva ML, Ventura D, Vielliard J, Mello CV 2000 Behaviourally driven gene expression reveals song nuclei in hummingbird brain. Nature 406(6796): 628–32
- Mayr G 2011 Metaves, Mirandornithes, Strisores and other novelties – a critical review of the higher-level phylogeny of neornithine birds. J Zool Syst Evol Res 49(1): 58–76
- Livezey BC, Zusi RL 2007 Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy: II. – Analysis and discussion. Zool J Linn Soc 149(1): 1–95
- Poe S, Chubb AL 2004 Birds in a bush: five genes indicate explosive evolution of avian orders. Evolution 58(2): 404–15
- Pratt RC, Gibb GC, Morgan-Richards M, Phillips MJ, Hendy MD, Penny D 2009 Toward resolving deep Neoaves phylogeny: data, signal enhancement, and priors. Mol Biol Evol 26: 313–26
- Rokas A, Carroll SB 2006 Bushes in the tree of life. PLoS Biol 4(11): e352
- Sibley CG, Ahlquist JE, Monroe BL Jr 1988 A classification of the living birds of the world based on DNA–DNA hybridization studies. Auk 105(3): 409–23
- Slack KE, Delsuc F, McLenachan PA, Arnason U, Penny D 2007 Resolving the root of the avian mitogenomic tree by breaking up long branches. Mol Phyl Evol 42: 1–13
- Sorenson MD, Oneal E, García-Moreno J, Mindell DP 2003 More taxa, more characters: the hoatzin problem is still unresolved. Mol Biol Evol 20: 1484–98
- Suh A, Paus M, Kiefmann M, Churakov G, Franke FA, Brosius J, Kriegs JO, Schmitz J 2011 Mesozoic retroposons reveal parrots as the closest living relatives of passerine birds. Nature Comms doi:10.1038/ncomms1448
- Waddell PJ, Kishino H, Ota R 2001 A phylogenetic foundation for comparative mammalian genomics. Genome Inform 12: 141–54
- Warren WC, Clayton DF, Ellegren H, Arnold AP, Hillier LW, Künstner A, Searle S, White S, Vilella AJ, Fairley S, Heger A, Kong L, Ponting CP, Jarvis ED, Mello CV, Minx P, Lovell P, Velho TA, Ferris M, Balakrishnan CN, Sinha S, Blatti C, London SE, Li Y, Lin YC, George J, Sweedler J, Southey B, Gunaratne P, Watson M, Nam K, Backström N, Smeds L, Nabholz B, Itoh Y, Whitney O, Pfenning AR, Howard J, Völker M, Skinner BM, Griffin DK, Ye L, McLaren WM, Flicek P, Quesada V, Velasco G, Lopez-Otin C, Puente XS, Olender T, Lancet D, Smit AF, Hubley R, Konkel MK, Walker JA, Batzer MA, Gu W, Pollock DD, Chen L, Cheng Z, Eichler EE, Stapley J, Slate J, Ekblom R, Birkhead T, Burke T, Burt D, Scharff C, Adam I, Richard H, Sultan M, Soldatov A, Lehrach H, Edwards SV, Yang SP, Li X, Graves T, Fulton L, Nelson J, Chinwalla A, Hou S, Mardis ER, Wilson RK 2010 The genome of a songbird. Nature 464(7289):757–62











0 komentářů:
Okomentovat
Povolené tagy:
- <b>tučně</b> = tučně
- <i>kurzíva</i> = kurzíva
- <a href="http://pan-aves.blogspot.com/">pan-Aves</a> = pan-Aves
Podporuje $\mathrm{\LaTeX}$ pro matematické vzorce.