
Tato recenze bude trochu netradiční. Nebudu se zde zabývat výhradně knihou samotnou (přestože by si to jistě zasloužila), spíše se budu soustředit na stavění jejích tvrzení do kontrastu s novějšími paleontologickými poznatky. Trvám na tom, že kniha velmi pěkně konzervuje stav, který v dinosauří paleontologii v dané éře (tj. na konci osmdesátých let) panoval, ale přijímat nekriticky její vývody dnes už nelze. Mnoho věcí zůstalo pravdou, mnoho věcí bylo vyvráceno,a některé z nich se ještě před pár lety zdály být vyvráceny, aby je dnes paleontologický konsenzus opět vzal zpět.
Nejprve se tedy musíme zaměřit na období, kdy byla kniha vydána a co pro něj bylo charakteristické, abychom ji mohli zhodnotit v náležitém kontextu. První vydání pochází z roku 1993. Při zpětném pohledu může tato doba vypadat z hlediska dinosauřího paleontologa poměrně nudně. Renesance, kterou obor prošel v sedmdesátých letech už byla pryč, a pohled na dinosaury jako endotermní, aktivní živočichy byl už poměrně mainstreamový (což ovšem platí pro tehdejší odbornou, nikoli laickou veřejnost - v jejím informovaní odvedla Záhada dinosaurů kus dobré práce). Kladistika, která naprosto změnila význam systematiky - ze které už nadále nebyl formalistický obor bez kontaktu s realitou, ale zásadní a často i jediný způsob pro odvozování anatomických a ekologických informací, které nemůžeme získat z fosilií - už svou první etapu měla také za sebou. Od Gauthierovy zásadní studie z roku 1986 uplynulo již sedm let, což v tak dynamickém odvětví není málo. Naopak do objevu opeřených dinosaurů z Číny, který obor dále zrevolucionizoval, několik let ještě zbývalo.
Jenže tak tomu není. Nevím, kdy J. Mareš začal knihu psát, je v ní však naprosto jasně patrné myšlenkové dědictví osmdesátých let. Ano, byla to sedmdesátá léta, kdy Ostrom obnovil sto let starou hypotézu o přímé evoluční souvislosti mezi dinosaury a ptáky, kdy byla dokázána dinosauří monofylie a kdy Bakker přinesl první evidenci o dinosauří endotermii. Ale v osmdesátých letech se objevilo ještě mnohem větší množství zajímavých myšlenek, často od heretických, spíše populárně-naučně než technicky píšících, zato ale neuvěřitelně předvídavých autorů. Důkazy o přímém evolučním vztahu mezi dinosaury a ptáky se dále prohlubovaly. Thulborn a Barsbold přinesli spolu s objevy nových, ptákům podobných teropodů důkazy o tom, že "prapták" Archaeopteryx je jen lehce odvozeným celurosaurem, nemajícím se vznikem ptáků mnoho společného. G. S. Paul, paleontolog samouk, maloval podle dosud nijak nepodložených Bakkerových hypotéz dinosaury s pernatým pokryvem těla a navrhl, že by některé linie teropodů mohly být ve skutečnosti druhotně nelétavými potomky ptáků. Stejně se rozvíjely i hypotézy o dinosauří endotermii. Systematika se začala měnit od základů - kromě návrhů o podstatném rozšíření obsahu Dinosauria se objevily i hypotézy o tom, že již jedno století rozeznávané taxony Saurischia a Ornithischia nemusí reprezentovat přesnou dinosauří dichotomii. Je zřejmé, že některé z divokých hypotéz, které se v tomto období objevily, nebyly pozdějšími výzkumy s přísnější metodologií potvrzeny.
Dinosauři, ptáci a obecná systematika
První věc, na kterou je třeba upozornit, je že "dinosauři", jak o nich píše J. Mareš, nejsou těmi samými dinosaury, jako ti, kteří plní stránky dnešních vědeckých publikací. Autor chápe pojem "dinosauři" daleko šířeji, než bylo v odborné literatuře běžné i v době prvního vydání Záhady. Kromě ptakopánvých a plazopánvých zde jsou mezi dinosaury řazeni i taxony jako Marasuchus, Lagosuchus a Lagerpeton, pterosauři a dokonce i malý "tekodont" Scleromochlus taylori, který je v některých fylogenetických hypotézách nejbazálnějším pan-avianem vůbec (Benton 1999). Marešovi "dinosauři" jsou tak názvem pro celou ptačí linii archosaurů, neboli ptačí total klad. V kontextu diskuzí o rozsahu Dinosauria v osmdesátých a na začátku devadesátých let se nejedná o nic neobvyklého. Bakker, který byl pro autora při psaní Záhady naprosto zjevnou a přiznanou inspirací, skutečně navrhl zahrnout pterosaury mezi dinosaury (Bakker 1986). Myšlenka, že pterosauři jsou velmi blízkými příbuznými tradičních dinosaurů již tehdy byla ve značné oblibě a potvrdily ji i kladistické analýzy. Gauthier (1984:175-182) dokonce pterosaury odhalil jako bližší příbuzné dinosaurům, než byl dinosauromorf Lagosuchus, což zjevně Bakkera inspirovalo. Jeho návrh byl tedy spíše formálním pokusem o stvrzení blízké příbuznosti pterosaurů s "tradičními" dinosaury. Takový způsob uvažování není nepodobný tomu, co J. Mareš vyjádřil v některých svých článcích:
Jestli je dinosaurus to, kde výrazně převládají dinosauří znaky nad znaky tekodontů, tak ptakoještěři jsou dinosauři. A pokud nasadíte jméno dinosaurus až tam, kde kompletně všechny znaky souhlasí s dinosaury, tak ptakoještěři mezi ně nepatří. Já se přikláním k tomu, že kde ty znaky převládají, tam už je možno mluvit o dinosaurech.
J. Mareš pro 21. Století (8/2003:83)
Zhruba na přelomu osmdesátých a devadesátých let však diskuze o obsahu Dinosauria skončila a aplikace tohoto jména se ustálila u méně obsáhlého taxonu. Bakkerův návrh nebyl přijat a systematika začala požadovat spíše komplexní výčet apomorfií, než méně přesný fenetický přístup operující s morfologickou a anatomickou vzdáleností různých skupin mezi sebou. Novas (1992a) formuloval první fylogenetickou definici Dinosauria, postavenou na herrerasauridech - taková definice by sice měla v některých tehdejších fylogenetických hypotézách větší obsah než klad Saurischia + Ornithischia (Eudinosauria sensu Novas), každopádně ovšem vylučovala marasucha, lagerpetona, pterosaury a skleromochla. Ještě D. B. Weishampel, P. Dodson a H. Osmólska ve svém úvodu k první edici The Dinosauria uvádějí, že hranice mezi non-dinosaury a dinosaury je subjektivní záležitostí a bylo by možné definovat jako dinosauří takové apomorfie, které jsou společné i pro pterosaury, ale že se postupně zformoval systematický konsezus, ve kterém pterosauři a Lagosuchus dinosaury nejsou, zatímco Herrerasaurus a Staurikosaurus ano (a to i navzdory tomu, že v dané fylogenetické hypotéze tyto taxony předcházely oddělení ptakopánvých a plazopánvých). Nelze však tvrdit, že by proti takovému pohledu neexistoval pozdější disent - "transformovaný kladista" Patterson (1993) navrhl, snad pod vlivem Bakkerovy interpretace, aby byl pojem Dinosauria kompletně opuštěn jako mladší synonymum Aves, které by se nyní aplikovalo na celý ptačí total klad!
Podobně široký obsah mají v Záhadě i savci. Snad kvůli lákavé představě prvních dinosaurů a savců, svádějících boje o ekologické niky se svými "tekodontními" a terapsidními předky a mezi sebou navzájem, byl jako savec uznán i Probainognathus, který údajně "je nyní již považován za prvního savce" kvůli svým silným a výrazným savčím adaptacím. Určitou logiku takový návrh má - Probainognathus je prvním známým synapsidem, který disponuje typickým skloubením čelisti za pomoci skvamosálu a zubní kosti, které je historicky užíváno jako významné kritérium pro určení savčí příslušnosti fosilií (pravda je však daleko komplikovanější a zajímavější: Probainognathus měl dvě funkční kloubení pro dolní čelist - jedno mezi skvamosálem a zubní kostí, apomorfické a "savčí", druhé mezi kostí čtvercovou a artikularem, pleziomorfické a typické pro všechny bazálnější amnioty). Z hlediska fylogenetických souvislostí by však bylo zahrnutí probainognáta mezi savce extrémním návrhem, vzhledem k pozici na kladogramu dost vzdálené dokonce i od bazálních mammaliaformů (na jejichž příslušnosti k Mammalia také nepanuje shoda) - v některých topologiích dokonce probainognáta od savců oddělují podle J. Mareše nesavčí tritylodonti (Luo et al. 2002). Pokud využijeme tradičnější definici Mammalia, vystihující paleontologické užívání jména (Luo et al. 2002), zjistíme, že první savci se objevili asi před 210 miliony let (kdy už za sebou dinosauři měli přes 20 milionů let nerušené kompetice s bazálnějšími archosaury); pokud se rozhodneme použít crown-clade definici, první savci vznikají až ve spodní juře, kdy už dinosauři bezpečně ovládají většinu ekosystémů.
Možná má však široký rozsah savců v Záhadě jiný důvod (tady už jenom spekuluji, nemám samozřejmě nejmenší ponětí, proč autor psal právě to, co psal). Během období, kdy mezi autory převažovala hypotéza o savčí polyfylii (té se J. Mareš drží - "Ale vznikem prvních savců jejich vývoj z terapsidů neskončil. Vznikali postupně z různých terapsidních větví a nejsou vůbec jednolitou skupinou." [str. 106]) - toto období se mimochodem pozoruhodně krylo s výskytem návrhů o polyfylii dinosaurů - se několikrát objevil návrh, aby byl taxon Mammalia učiněn monofyletickým tím, že do něj bude zahrnuto i množství primitivnějších synapsidů tak, aby taxon obsahoval společného předka jednotlivých synapsidních větví, které měly stát za vznikem různých skupin savců (Simpson 1959; Reed 1960 [Luo et al. 2002]). Problém je v tom, že koncept polyfyletických savců byl mezi paleontology zavržen již v sedmdesátých letech, sotva pár let před tím, než Bakker a Galton oživili monofyletické dinosaury (Luo et al. 2002). Měla snad Záhada dinosaurů v době svého vydání dvacetileté zpoždění, co se původu savců týče?
Zřejmě ne. Jako mnoho jiných kontroverzních tvrzení v knize, i to o raných savcích lze dohledat v Paulově knize Predatory Dinosaurs of the World:
A popular misconception is that mammals appeared much later than the first dinosaurs. Actually, protomammals appear the same time as protodinosaurs, at about 235 million years ago in the mid-Triassic, and the two groups shared the rest of the Mesozoic Era.
Upřímně by mě zajímalo, kde tyto informace G. S. Paul vzal - tento autor je však znám tím, že podává autoritativním tónem i informace, o kterých vůbec jistota nepanuje.
Kromě nezvykle velkého obsahu savců je líčení nástupu dinosaurů ve střední triasu skvělým textem, téměř ve vojenském stylu, kterému lze málo co vytknout. Autor rozeznává dvě hlavní skupiny triasových amniot: terapsidy a "tekodonty". Obě jsou zde používány, vzhledem ke své době nijak překvapivě nebo nesprávně, v parafyletickém významu - živočich tedy může být buď terapsidem, nebo savcem, ne však obojím najednou. "Tekodonti", dnes už nepoužívaný parafyletický grad bazálních archosauriformů, je dělen do tří skupin - proterosuchianů, aetosaurů a pseudosuchianů. Tradiční klasifikace zahrnuje ještě čtvrtou skupinu - Phytosauria - kterou J. Mareš také zmiňuje, ne však v taxonomických souvislostech. Trochu překvapivě J. Mareš začíná úspěch dinosaurů už ve svrchním permu u terapsidů, jejichž nadřazenost vůči dosud existujícím plazím skupinám popisuje za pomoci porovnávání s dnešními savci. Přes nástup tekodontů se však rychle dostáváme k prvním dinosaurům. Již jsme zjistili, že v Záhadě je tento pojem chápán podstatně šířeji než dnes, a sám autor uvádí, že kritéria "jsou u různých autorů odlišná". Přestože J. Mareš píše, že "postupně [...] převládl názor," je zřejmé, že inspirací je zde opět Paul, už jen z používání pojmu "protodinosauři". Jako první dinosauři jsou tedy uváděni dinosauromorfové Lagerpeton canarensis, Lagosuchus talampayensis, Pseudolagosuchus major a s ním pravděpodobně synonymní Lewisuchus admixtus (Arcucci 1998). "Lagosuchus" lilloensis, kterého J. Mareš uvádí jako mladší synonymum L. talampayensis, si paradoxně později vysloužil i generickou separaci pod jménem Marasuchus (Sereno & Arcucci 1994). Výčet karnských dinosaurů je na tom o něco lépe - jejich dinosauří status zůstal nezměněn, ale dva rody a druhy, které J. Mareš (a potažmo Paul) rozeznává - Frenguellisaurus ischigualastensis a Ischisaurus cattoi - se ukázaly být pouhými synonymy H. ischigualastensis (Novas 1994). Aliwalia rex - domnělý gigantický, 8 m dlouhý jihoafrický herrerasaurid (Galton 1985; Paul 1988a) - je zřejmě bazálním sauropodomorfem z kladu Riojasauridae a synonymem druhu Eucnemesaurus fortis. S tím padá lákavá představa predátorů velikosti allosaura, vznikajících už 10 milionů let po samotném úsvitu dinosaurů. Ten je v knize datován do období před 235 miliony, ke kterému lze i přes podstatně širší obsažnost dinosaurů v Záhadě stěží mít námitky, a to i přesto, že nejstarší dobře zdokumentovaný dinosauří materiál pochází z velmi raného karnu asi před 230 miliony let (Langer et al. 2009). Vzhledem k tomu, že už v raném karnu se dinosauři zdají být rozrůzněni do relativně velkého počtu linií (Ornithischia [Pisanosaurus], Herrerasauria [Staurikosaurus, Herrerasaurus], Sauropodomorpha [Panphagia, Saturnalia] a ?Theropoda [Eoraptor]), a navrhované sesterské taxony dinosaurů (Marasuchus, Pseudolagosuchus, klad [Silesaurus + Pseudolagosuchus + Lewisuchus]) jsou nalézány v ladinských, střednětriasových vrstvách, zdá se být ladinský původ dinosaurů pravděpodobný (Langer et al. 2009). Martinez & Alcobar (2009) dokonce navrhli, že ve středním triasu mohlo dojít nejen ke vzniku, ale i základní diverzifikaci dinosaurů včetně vzniku sauropodomorfů, což je konec konců podpořeno možným pozdně ladinským stařím saturnalie (Langer 2004).
Oproti tomu líčení savčího osudu po dinosauřím "vítězství" je již neaktuální i přinejmenším nepřesné. Představa savců jako drobných rejskovitých zvířátek, které dinosauři na desítky milionů let odsoudili k životu v úkrytech, je jistě velmi přitažlivá, má však mnoho trhlin. Druhohorní savce větší než 30 centimetrů známe - patří mezi ně např. dravý vačnatec Didelphodon, metrový spodnokřídový ptakořitný Kollikodon, multituberkulát Bubodens a samozřejmě trikonodont Repenomamus, jehož kořistí se stávali i dinosauři (konkrétně malý Psittacosaurus). Molekulárně biologické studie ukazují, že savci nezaznamenali svůj první úspěch až s oportunistickým využitím ekologických nik uvolněných vyhynutím non-avianních dinosaurů na přelomu křídy a paleocénu, ale prodělali své hlavní radiace během koexistence s dinosaury. Pomineme-li fakt, že k samotnému vzniku savců došlo za existence dinosaurů, k odštěpení ptakořitných došlo ve střední juře, k oddělení vačnatců od plancentálů v pozdní juře, a k velké eutherianní radiaci, při níž vznikly všechny dnešní velké skupiny placentálů, došlo v intervalu mezi 100 - 74 miliony let ode dneška (Bininda-Emonds et al. 2007). To znamená, že "zlatý věk" pro dinosaury a savce, alespoň z hlediska diverzifikace, se prakticky překrýval - byla jím pozdní svrchní křída.
Ptáci a jejich původ
Ptáci a jejich původ
Pro čtenáře obeznámeného s vývojem představ o dinosaurech, ptácích a jejich vzájemném vztahu bude čtení Záhady lahůdkou. Vezměme si např. následující úryvek:
Kladistika a systematika dinosaurů
Ilustrace
Ke knize
Zdroje:
Podklady:
Dlouhá snaha mnoha paleontologů najít prvního ptáka nemá vlastně smysl a je ekvivalentní příslovečnému hledání pramenů Nilu. Jako Nil, vytékající z Viktoriina jezera, je tvořen množstvím pramenů, tak i ptáci, jak je známé dnes, nejsou a nikdy nebyli jednolitou skupinou, ale společné charakteristiky u nich vznikly konvergentním vývojem několika skupin dravých dinosaurů, kteří díky opeření získali schopnost letu každá zvlášť a v jiné době.
Zde se naprosto zřetelně odrážejí Barsboldovy ideje o "ornitizaci", postupném, masivně konvergentním procesu získávání ptačích znaků u odvozených teropodů. Zatímco dnes je obecně přijímano jako fakt, že většina podobností mezi coelurosaury a ptáky je homologické povahy, Barsboldova ornitizace je stále dobrým popisem tendencí některých skupin vyvíjet znaky podobné těm, které nacházíme až u daleko odvozenějších ptáků. Tyto tendence mohou být obecnější (např. zkrácení kaudální série u bazálních oviraptorosaurů) nebo velmi úzce ohraničené (karpometakarpus avimima, ztužení distální části ocasu do jakéhosi "protopygostylu" u nomingie a similikaudipteryga).
Odkud se bere Marešův návrh, podle kterého by Avimimus mohl být předkem evoluční linie vedoucí k dnešním pštrosům, čili ratitů? Barsbold (1983b) ve své práci o avimimovi sice nenavrhl přímo takovou souvislost, ale navrhl polyfylii žijících ptáků, kde ratitní a karinátní nesdílejí společného ptačího předka. Takový návrh je v moderní literatuře a také celé historii výzkumu ptačího původu poměrně unikátní. Občas sice byla zpochybněna ptačí příslušnost určitých taxonů známých z fosilního záznamu - Archaeopteryx je typickým příkladem - ale monofylie žijících ptáků jako takových byla napadnuta jen málokdy, vzhledem k obrovskému množství sjednocujících znaků. Pokud je mi známo, v odborné literatuře nikdy přímá vývojová linie vedoucí od avimima k ratitům navržena nebyla, ale Záhada dinosaurů je populárně-naučného zaměření a autor si mohl dovolit spekulovat podle libosti. Avimimus je pak tip stejně dobrý jako kterýkoli jiný - ve skutečnosti lepší než většina jiných, díky svému karpometakarpu, domnělé absenci ocasu (zjevné na rekonstrukci B. Kyškové) a přítomnosti peří, předpokládané kvůli strukturám podobným "quill knobs" na kosti loketní (Kurzanov 1987).
Zajímavou, byť v kontextu nevýznamnou narážkou na tehdejší fylogenetické hypotézy je i polyfylie letounů (Chiroptera). Podle Záhady se netopýři vyvinuli z hmyzožravců, zatímco kaloni z plodožravých opic. Pettigrew (1986; Pettigrew et al. 1989) skutečně navrhoval, že megachiropterani ve skutečnosti jsou létajícími primáty, a pro podporu této teorie předložil několik komplexních znaků vizuálního systému, molekulární biologie a kladistika však poměrně záhy tuto hypotézu smetla (Simmons 1994), podobně jako polyfylii žijících ptáků.
Jako další příklad překonaných poznatků, které Záhada nabízí, je možné vzít zde uváděný model vývoje peří:
Jak jsme si již řekli, museli si endotermní dinosauři žijící v chladnějších oblastech a především jejich menší formy chránit svou tělesnou teplotu nějakým tělesným pokryvem. Východiskem byla přeměna šupin ve zřasený, výborně izolující útvar. Vznikla primitivní forma péra. Šupiny se zřejmě nejdříve listovitě prodloužily a překrývaly se, čímž tepelnou izolaci zlepšily. Každá z nich byla podpírána středovým žebrem s postranními větvemi. Dalším stadiem bylo pravděpodobně oddělení těchto větví do samostatných "lístků" a nakonec se tenká blána mezi nimi roztřepila v mikroskopickém měřítku na praporky a paprsky, které vytvářejí stavbu péra dnes.
Tento model zní naprosto logicky, paradoxně je však více kompatibilní s "tekodontní" než dinosauří hypotézou původu ptáků, kterou J. Mareš pomáhá popularizovat - vzhledem k tomu, že od dinosaurů šupiny v pravém smyslu slova (= vzájemně se překrývající epidermální struktury složené z keratinu) neznáme a nemáme důvod předpokládat, že jimi kdy tato skupina disponovala. Prum (1999; Prum & Brush 2002) na základě embryologických dat prokázal, že peří se nevytváří rozkladem již existujícího, komplexního epidermálního útvaru (tedy šupiny), ale vznikají z pokožky jako naprosto samostatná struktura, procházející komplikovaným vývojem od jediného vlákna bez folikulu a větví až po složitou strukturu s výrazným ostnem a kompaktním praporem. Prumova teorie z oblasti evolučně developmentární biologie má daleko lepší podklad ve fosilním záznamu než předchozí hypotézy: zatímco existuje dobrá evidence pro různá stadia vývoje peří od jediného vlákna až po různé stupně větvení (Xu & Guo 2009), neexistuje žádný doklad pro čím dál tím jemněji se řasící šupiny.
Jiným z atraktivních, nicméně již překonaných scénářů, které Záhada nabízí, je triasová "slepá ulička" ve vývoji ptáků, Chatterjeeův Protoavis texensis. Úsek popisující protoavise není dlouhý a autor v něm předkládá pouze základní fakta, interpretovaná v rámci Barsboldovy ornitizace. Nejsou zde uvedeny Paulovy spekulace o silné konvergenci odvozených herrerasauridů s ptáky, stvrzené pojmenováním Herreravia. (Afinita spatřovaná Paulem mezi protoavisem a herrerasauridy je však založena pouze na faktu, že jde o jediné teropody s dostatečně brzkým stratigrafickým výskytem na to, aby se mohlo jednat o protoavisovy předky.) Přestože mnoho paleontologů bylo k posunutí ptačího původu do triasu skeptických už od začátku (Ostrom 1987, 1991; Wellnhofer 1992), detailní kritika Chatterjeeových závěrů, argumentující chimérickým statutem protoavisových fosilií, se objevila až později, po vydání Záhady.
Archaeopteryx je poměrně neobvykle označen za "mal[ého] teropod[a] s výraznými ptačími znaky" místo tradičnějších přízvisek typu "první pták". J. Mareš rovněž uvádí, že i přes tato častá označení byl Archaeopteryx "zástupcem slepé větve, která k vývoji dnešních ptáků nevedla". Vzhledem k tomu, že jednou z hlavních autorových inspirací byla Paulova kniha Predatory Dinosaurs of the World (Paul 1988a), není něco takového překvapivé. V této polotechnické publikaci, která měla spolu s Bakkerovými The Dinosaur Heresies na Záhadu značný vliv, autor na základě komparativní anatomie navrhoval, že by některé historicky neptačí linie teropodů mohli mít vlastně blíže k žijícím ptákům, než Archaeopteryx, a být druhotně nelétavé. G. S. Paul není první, kdo s tímto návrhem přišel - Thulborn publikoval na stejné téma celou sérii článků, od svého revolučního návrhu, aby se jméno Aves aplikovalo na všechny teropody (Thulborn 1975), až po kladistickou analýzu, ve které díky ne zcela informativním znakům odkryl takovou topologii coelurosaurů, ve které jsou nejen Avimimus a troodontidi, ale dokonce i tyrannosauridi (!) blíže příbuzní žijícím ptákům, než Archaeopteryx nebo dromaeosauridi (Thulborn 1984). Logickým závěrem tedy bylo, že Archaeopteryx nejenže nestál u počátků ptačí evoluce, jak se domníval Ostrom a vlastně i většina autorů od 19. století, ale že by ani neměl být nazýván ptákem (ve smyslu Aves), neboť by něco takového muselo nevyhnutelně vést i k zahrnutí mnoha dalších teropodích linií. Přestože Paul stejný postoj nepřestal obhajovat až do současnosti (Paul 2002) a některé dnešní výzkumy rozložení maniraptořích synapomorfií mu dávají za pravdu v tom, že nejen peří, ale i let v dinosauří evoluci archaeopteryga předcházel (Longrich 2006), kladistické analýzy, počínaje tou Gauthierovou z roku 1986, úzký vztah archaeopteryga s žijícími ptáky - vylučující všechny historicky neptačí teropody - nevyvrátily. Podobně jako v části o významu archaeopteryga pro ptačí evoluci, i informace o jeho ekologii jsou poněkud neaktuální nebo lépe řečeno sporné. Debaty o tom, zda Archaeopteryx mohl aktivně létat nebo ne sahají dlouho do minulosti, jelikož mají značný význam pro rozhodování mezi "ground-up" a "trees-down" scénáři vzniku letu. J. Mareš na základě blíže neurčených posledních studií píše, že ano - v současnosti se však jeví jako pravděpodobnější opačná možnost. Z anatomických dat bylo odvozeno, že pro archaeopteryga bylo biomechanicky nemožné zvednout paže nad úroveň dorzální páteře - jeho lopatky měly glenoid, jamku pro skloubení s humerem, orientovanou ventrálně místo laterálně (Senter 2006). To dokazuje, že Archaeopteryx byl nejen neschopen aktivního letu, ale zřejmě také těžko mohl běhat po písečných plážích, vyskakovat a lapat do svých křídel hmyz, jak J. Mareš popisuje ground-up scénář. Bakkerovy spekulace o potápějícím se archaeopterygovi jsou takto uvedeny a lze je považovat spíše za předloženou zajímavost.
Velký význam pro poznání raných fází evoluce ptáků autor přikládá dosud nepojmenovanému "praptákovi" z Liaoningu, "nazývan[ému] předběžným jménem sinopteryx". Má být jen o 10 milionů let mladší než Archaeopteryx, zato však již s moderními ptačími znaky včetně pygostylu nebo osifikovaného sterna s masivním kýlem, zabezpečujícím schopnost aktivního letu. O který taxon jde? Je jím Sinornis santensis (viz Sereno et al. 2002), který skutečně pygostylem a schopností aktivního letu disponoval, už jen příslušnost k Enantiornithes - které sám J. Mareš uvádí jako vývojovou větev bez spojitosti s žijícími ptáky - dává vědět, že praotcem dnešních ptáků zřejmě nebude. Sinornis rovněž není starý 140 milionů let - přestože je datování Jiufotang Formation, ve které byl nalezen, poněkud problematické, už z prostého faktu, že její vrstvy překrývají souvrství Yixian a Chaomidianzi barremského stáří (tj. kolem 130 - 120 Mya), se dá určit nejspíše albský nebo aptský původ (tedy 125 - 99,6 Mya).
Kladistika a systematika dinosaurů
A co kladistika? I když v roce 1993, kdy byla Záhada vydána, už lze kladistiku v dinosauří paleontologii těžko označit jako nastupující - zásadní analýzy, jako ty v pracích Padian 1982; Gauthier & Padian 1985; Gauthier 1986 už byly provedeny - její aplikace stále nebyla tak masivní jako dnes. Když jsem se po dvou letech opět do Záhady začetl, jaké bylo mé překvapení, když jsem o ní našel jedinou, avšak přímou zmínku:
"Kladistický rozbor základních znaků jasně řadí ptáky do třídy Dinosauria," prohlásil v roce 1991 jeden z nejvýznačnějších světových paleontologů, Kanaďan Philip J. Currie, specializující se právě na malé teropodní dinosaury a vznik prvních ptáků.
Nepřímých zmínek je ovšem v knize daleko více. "Jacques Gauthier [...] zjistil přes 120 znaků, které jsou společné pouze dinosaurům a ptákům [...]" - zde se jedná o odkaz na studii Gauthier & Padian 1985, která obsahovala jednu z prvních kladistických analýz s (posuzováno dnešními měřítky) dostatečně velkou datovou matricí.
V knize (přinejmenším v některých jejích částech) je rovněž patrný důraz na monofylii taxonů, jdoucí ruku v ruce s rozšiřujícím se uplatněním kladistiky - a to dokonce i přesto, že autor se podle svých vlastních slov snažil diskuzím o systematice vyhýbat. Myšlenka, že pokud je nějaký taxon potomkem dinosaurů, měl by být i jejich zástupcem, je např. vyjádřena v kapitole "Létající dinosauři":
Esovitě zahnutý krk a další kosterní charakteristiky v oblasti ramen a kotníků těchto rychlých a hbitých prvních triasových dinosaurů napovídají, že právě oni dali vznik pterosaurům, prvním obratlovcům, kteří kdy dobyli vzduch aktivním letem. Stoupenci této teorie jsou i další paleontologové, jako např. Bakker, Currie aj. V tom případě by měli být ovšem i ptakoještěři proti všem vžitým dřívějším představám zařazeni mezi dinosaury.
Příkladů lze ovšem po celé knize najít daleko víc. Autor sice celkem pochopitelně nepoužívá pojmy jako monofylie, parafylie apod., rozlišuje ovšem "přirozené" a "uměle vytvořené" skupiny. Typickou ukázkou je např. kapitola o triasových dinosaurech a zvláště prosauropodech. J. Mareš v ní mimo jiné reviduje i názory na dinosauří polyfylii, provázejících paleontologii po většinu 20. století až do prací Padiana, Bakkera a Galtona. Na tomto místě uvádí, že dinosauři ze skupiny Ornithischia jsou potomky skupiny Saurischia, konkrétně anchisaurům podobných prosauropodů, a že "všichni býložraví dinosauři tvoří jednu přirozenou skupinu", nazvanou zde Bakkerovým jménem Phytodinosauria. Návrhy na to, že v rámci monofyletických dinosaurů existuje základní rozdělení na klady (Sauropodomorpha + Ornithischia) a Theropoda spíše než na (Sauropodomorpha + Theropoda) a Ornithischia, se objevily uprostřed osmdesátých letech a nikdy se nesetkaly s velkou podporou; kladistické analýzy včetně té mimořádně vlivné Gauthierovy je nikdy nevykryly. Cooper (1985) použil pro klad tvořený sauropodomorfy a ptakopánvými a zároveň vylučující teropody jméno Ornithischiformes. Toto neobvyklé dělení bylo v jeho práci doprovázeno neobvyklou fylogenetickou strukturou ptakopánvých a užíváním téměř zapomenutých nebo zcela nových jmen pro dobře známé taxony (Pachypodosauria pro Sauropodomorpha, Carnosauriformes pro Theropoda). Se stejnou hypotézou přišel nezávisle na Cooperovi o rok později Bakker (1986), který domnělý klad non-teropodních dinosaurů nazval Phytodinosauria. Nesetkal se však s širší podporou systematiků a v době vydání Záhady už neměla fytodinosauří hypotéza v diskuzi o dinosauří fylogenezi žádný význam. Její zahrnutí do knihy a předložení čtenářům jako fakt je tedy poněkud překvapivé - už v době publikace byla její podpora značně limitovaná, narozdíl od témat, která s ní J. Mareš dává do souvislosti (např. dinosauří monofylie). Na druhou stranu není neočekávané, jelikož se ke stejné hypotéze přikláněl i Paul - v Predatory Dinosaurs of the World zmiňuje fytodinosaury jako jednu z hlavních dinosauřích skupin, spolu s teropody, nejistě umístěnými herreraviany a primitivními paleodinosaury, a ve své dřívější práci dokonce uvádí segnosaury (= terizinosaury) jako pozůstatky přechodu mezi prosauropody a ptakopánvými (Paul 1984a). Z čeho vyplývá argumentace pro monofylii fytodinosaurů?
Alan Charig noted back in 1976 that if dinosaurs are monophyletic, then Saurischia and Ornithischia are no longer equivalent clades. Saurischians are just those dinosaurs that happen to retain relatively primitive hips, and ornithischians are just one of many groups that evolved "bird hips."
(Paul 1988a:185)
Proti takové argumentaci lze stěží něco namítat. Opistopubická pánev byla skutečně vyvinuta v průběhu dinosauří evoluce vícekrát po sobě (u ptakopánvých, maniraptorů a také u herrerasaura), ptakopánví dinosauři jistě pocházejí z plazopánvých předků a dá se očekávat, že ancestrálním stavem dinosaurů byla propubická pánev (Na druhou stranu, vzhledem k tomu, že Herrerasaurus, jeden z nejprimitivnějších plazopánvých, disponuje opistopubickou pánví, stejně jako ptakopánví [ačkoli opistopubie u pisanosaura, vůbec nejbazálnějšího ptakopánvého, nemůže být potvrzena - viz Norman et al. 2004], mohlo by se snad jednat o synapomorfii - to by však musela propubická pánev nejen bazálních teropodů a sauropodomorfů, ale i eoraptora a stautikosaura reprezentovat druhotný zvrat). Kladistika ovšem s Paulovými a Bakkerovými závěry zamíchala, když nejen nepotvrdila, že ptakopánví jsou potomky (a tudíž i součástí) prosauropodů, ale také když našla přesvědčivé množství apomorfií pro saurischiany, skupinu původně postavenou na plesiomorfickém stavu pánve. Připočítat je nutné i fakt, že fylogenetická nomenklatura (přesněji Padian a May pět let po Paulově knize) dala jménům Saurischia a Ornithischia definici, která je skutečně ekvivalentními klady učinila (Padian & May 1993).
Jednou z největších změn od dob sepsání Záhady určitě prošla systematika sauropodů; nesetkáme se zde s žádným z pojmů, které jsou nám povědomé z dnešních prací a naopak. Jelikož se autor snažil - v populárně-naučné publikaci jistě rozumně - vyhnout řešení taxonomických otázek, místo s formálními jmény se v knize setkáme se skupinami, jejichž vymezení má spíše ekologický než fylogenetický význam; souvisí konkrétně se zdrojem potravy. Teď mám samozřejmě na mysli dělení sauropodů na ty s robustními (lopatkovitými) zuby, vhodné pro spásání tužší potravy, např. listů na stromech, a ty s gracilními (kolíkovitými) zuby vhodnými pro křehčí vegetaci. Obě skupiny velmi hrubě odpovídají kladům Diplodocoidea a Macronaria - zatímco diplodokidi a dikreosauridi měli jemnější typ zubů, kamarasauridi a brachiosauridi (v obou případech bazální makronariani) disponovali těmi lopatkovitými. J. Mareš správně uvádí, že lopatkovité zuby měli i křídoví "titanosauridi", píše však, že ti vyvinuli tento typ zubů nezávisle na brachiosauridech, což však nemusí být pravda - obě skupiny jsou si blízce příbuzné a jimi tvořený monofyl vylučuje taxony s kolíkovitými zuby. Na druhou stranu však robustním typem chrupu byli vybaveni i "mamenchisauridi", což je skupina, do které kromě mamenchisaura samotného může (Allain & Aquesbi 2008) a nemusí (Wilson 2002) spadat ještě Omeisaurus, a která stojí mimo - avšak velmi blízko od - neosauropodů (klad tvořený dikreosauridy, diplodokidy, brachiosauridy a "kamarasauridy"). Obzvláště problematická je pak skupina Cetiosauridae, která je naprosto typickou ukázkou "wastebasket taxonu", do kterého je zahrnováno vše, co nejde zařadit nějak lépe. Zajímavé je, že na jiných místech J. Mareš tento pre-kladistický taxonomický zvyk kritizuje (nebo přinejmenším zmiňuje jako překonaný), ale cetiosauridy zachovává - přitom by určitě popisu jednotlivých sem řazených sauropodů neublížilo, kdyby byla taxonomická informace zcela vynechána. Mezi cetiosauridy je v knize řazen mimojiné Haplocanthosaurus, na jehož příbuzenství existuje skutečně mnoho názorů (může se jednat o non-neosauropodního sauropoda [Upchurch 1995, 1998], např. blízkého příbuzného kamarasaura a brachiosauridů [Upchurch 1995]; velmi bazálního makronariana [Wilson & Sereno 1998]; nebo non-diplodokimorfního diplodokoida [Calvo & Salgado 1995; Wilson 2002]). Dále sem měl spadat Shunosaurus (bazální eusauropod, v některých méně podložených fylogenetických hypotézách tvořící součást monofyletických "euhelopodidů") nebo Chinshakiangosaurus, který skutečně velice bazálním sauropodem je (Upchurch et al. 2007b). Vulcanodon je uváděn jako "přechodný stupeň mezi prosauropody a sauropody" a je naznačeno, že jeho sauropodní pozice není jistá. Je pravda, že pozice tohoto taxonu mezi sauropody byla vždy okrajová (Salgado et al. 1997 jej dokonce využili při prvním pokusu fylogeneticky definovat sauropody jako jednoho ze specifikátorů), s využitím Serenovy branch-based definice se však v moderní systematice mezi sauropody pohodlně vejde nejen Vulcanodon, ale i množství dalších taxonů včetně několika linií historických prosauropodů (záleží na fylogenetické hypotéze - např. v Smith & Pol 2007 Sauropoda zahrnuje i taxony Gongxianosaurus, Camelotia, Antetonitrus, Lessemsaurus, Anchisauridae, "Blikanasauridae", "Melanorosauridae" a "Yunnanosauridae"). Pokud jde o cetiosauridní příslušnost breviparopa, zde ani nemá cenu tuto spekulaci komentovat. Cetiosauridae v Záhadě je buďto extrémně polyfyletickou, nebo parafyletickou skupinou rozptýlenou po celém kladu Sauropoda.
Ani u teropodí systematiky se nesetkáme se známými pojmy typu "tetanura" nebo "maniraptor", na druhou stranu autorovi slouží ke cti, že vynechal i Bakkerovy ("Dinoaves") nebo Paulovy ("Protoavia") ekvivalenty těchto jmen. K fylogenezi načrtnuté u teropodů lze mít z dnešního pohledu podstatně méně výhrad - Dilophosaurus je např. uváděn jako možný přímý potomek coelophysise. Hledání přímých vztahů "předek-potomek" je sice v kladistice neobvyklé, některé nové kladogramy však naznačenou interpretaci skutečně připouštějí. Jiné detaily sice příliš neodpovídají: Gasosaurus a Piatnitzkysaurus jsou - jako nejbazálnější tetanura vůbec v prvním případě a bazální megalosauroid v případě druhém - ve skutečnosti vývojově méně pokročilí než "megalosauridi". Typickou ukázkou je také reprodukování Paulova návrhu, podle nějž jsou tyrannosauridi přímými potomky allosauridů, což by v kladistickém pohledu znamenalo, že druhá skupina zahrnuje tu první. Nechybí dokonce ani komentáře, podle nichž jsou např. Carcharodontosaurus saharicus (non-allosauridní allosauroid) a Bahariasaurus ingens (který může být jak karcharodontosauridním karnosaurem [Rauhut 1995], tak bazálním coelurosaurem, jak naznačuje kladogram Andrey Caua) "velmi pokročilí allosauři stojící už těsně na hranici přechodu do čeledi Tyrannosauridae" - zdrojem jsou nečekaně Predatory Dinosaurs of the World (str. 317). Myšlenka, že tyrannosauři jsou skutečně karnosauři, a tedy bližší příbuzní jiných robustně stavěných teropodů než malých agilních dravců z kladu Coelurosauria, se těšila určité podpoře ještě v začátcích kladistiky a byla jí i podložena (Gauthier 1986; Bonaparte et al. 1990; Molnar et al. 1990), brzy se ale ukázalo, že spíše opak je pravdou. Paulova fylogeneze teropodů z roku 1988 se dnes vůbec zdá být v mnoha věcech jednoduše "mimo" - např. návrh na blízkou příbuznost allosauridů, druhu Ornitholestes hermanni (bazální maniraptor), piveteausaura (?bazální tetanury) a proceratosaura (snad tyrannosauroida [Rauhut et al. 2009]). A ještě jedna velice zajímavá hypotézu, zmíněná v Záhadě, si zaslouží podrobnější náhled. Jde o interpretaci "segnosaurů" (= klad Therizinosauria sensu Sereno) jako zbytku po vývojovém posunu od prosauropodů k ptakopánvým, tedy v souladu s fytodinosauří hypotézou. J. Mareš správně uvádí, že tato skupina disponuje ptačí pánví - to ale není nic neobvyklého ani u pokročilých teropodů, jakými jsou alvarezsauridi, dromaeosauridi, přinejmenším bazální troodontidi a ptáci; zdá se, že u oviraptorosaurů došlo k druhotnému zvratu od ancestrálního maniraptořího stavu, kterým skutečně byla opistopubická pánev [Holtz & Osmólska 2004]). Interpretace, že segnosauři, známí v době vydání Záhady pouze z křídových sedimentů, jsou pozůstatky hypotetické spodnojurské kladogenetické události - odštěpení se nové ptakopánvé linie od prosauropodů - bylo velmi odvážné a špatně podložené. Zatímco kladistika ukázala, že terizinosauři jsou odvození maniraptorní teropodi a jeden z mnoha dokladů vývojové tendence teropodů k býložravosti, z Číny byl skutečně popsán "segnosaur" spodnojurského stáří - Eshanosaurus (Xu et al. 2001). Paleontologové byli k terizinosauří identitě tohoto taxonu skeptičtí - jednak proto, že vyžaduje extrémně dlouhé "ghost lineages" u většiny celurosauřích kladů, jednak z toho důvodu, že v kladistických analýzách vycházel jako odvozenější než spodnokřídový Falcarius. Barrett (2009) však předložil důkazy pro správnost původní interpretace eshanosaura. Není však třeba dovolávat se prosauropodního původu terizinosaurů - objevy z tohoto roku ukázaly, že netušená evoluční historie se skrývá za mnoha dalšími maniraptořími klady. Původní kvadrupední, prosauropodům podobné rekonstrukce, jako ta ze strany 215, jsou dnes již zastaralé. A ano, samozřejmě, ještě jeden detail: chybí jim peří.
Velký, větší, největší
Záhada dinosaurů je také oblíbeným zdrojem pro nejrůznější žebříčky dinosauřích rekordů, přestože v tomto ohledu je její neaktuálnost rozeznatelná snad nejsnadněji. Jde o kvalitní knihu a autor se nemusel pro získání pozornosti čtenářů uchýlit k minimům a maximům vytrženým z kontextu, což se nedá říct o mnoha jiných, "populárně-naučných" knihách o dinosaurech. Na tom, že údaje Záhady jsou dnes přinejlepším nepřesné, autor ve většině případů nenese vinu. Mnoho fosilií, o kterých J. Mareš píše, bylo později reinterpretováno. Obrovská délka "seismosaura" byla zřejmě pouze výsledkem špatné identifikace kaudálních obratlů, která ovlivnila domnělou délku ocasu a proporčně tedy celého těla (Lucas et al. 2004). "Ultrasaurus" je chimérou, složenou z pozůstatků druhů Brachiosaurus altithorax a Supersaurus vivianae, a nejedná se tedy o 130 tun těžkého, jednadvacet metrů vysokého brachiosaurida. Většina hmotnostních odhadů pro sauropody v knize je již beznadějně zastaralá - důvody pro to však nejsou úplně jasné. Je samozřejmé, že výzkum pneumatizace sauropodomorfní kostry od konce osmdesátých let značně pokročil, ale i před tím došlo mnoho autorů k odhadům, které se zdají být i z dnešního hlediska přesné a realistické. Proč se např. J. Mareš rozhodl citovat zastaralé, 80-ti nebo dokonce 100-tunové odhady pro brachiosaura (Kurtén 1968), když práce Paula (1988b) na základě přesnějších výpočtů a lepší evidence nastínila hmotnost ani ne poloviční a k podobným výsledkům došli i jiní autoři? Důvodem mohl být samozřejmě i prostý fakt, že J. Mareš nebyl s Paulovou prací seznámen - něco, co lze těžko posoudit (citovaná literatura zahrnuje pouze knihy, ne články z žurnálů). Podložený není ani 33 metrový odhad délky pro druh Brachiosaurus brancai (resp. Giraffatitan brancai). Jaký materiál má přesně J. Mareš na mysli, když zmiňuje tendagurské "ojedinělé kosti enormních rozměrů"? Zřejmě mezi ně patří fibula HMN XV2. Ta je ale se svou délkou 1340 mm pouze o 13% delší než ta od téměř kompletního, v Berlíně vystaveného exempláře HMN SII ustanoveného Taylorem (2009) jako lektotyp druhu Giraffatitan brancai, která měří 1190 mm (Paul 1988b:3). Vzhledem k tomu, že délka HMN SII se pohybuje kolem 22 metrů (22,46 podle Janensche [1950b], 22,2 podle Paula [1988b]), extrapolací získáme délku 25,1 metru pro HMN XV2 ("25 estimated" podle Paula). Vzhledem k tomu, že HMN SII je pro svůj ne zcela srostlý skapulakorakoid zřejmě subadultní jedinec, vypovídá údaj vyvozený z HMN XV2 o délce dospělého giraffatitana možná i lépe. Paul (1988b) připomíná, že ani další tendagurské fosilie přisuzované extrémně velkým brachiosaurům nemusely ve skutečnosti patřit zvlášť gigantickým zástupcům: skapulakorakoid BYU 5001 byl oproti skapulakorakoidu exempláře HMN SII zhruba o tolik větší, o kolik byla fibula druhého z nich menší oproti HMN XV2. Tím se opět dostáváme k délce asi 25 metrů (Paul 1988b:3). Tím pádem ani brachiosauridní část "ultrasaurova" materiálu není dostatečně velká, aby z ní šlo vyvozovat rozměry jako 33 metrů délky nebo 21 metrů výšky - skapulakorakoid BYU 9462 není větší než tendagurský materiál patřící giraffatitanovi (Taylor 2009). Ale ačkoli izolované tendagurské kosti neukazují na brachiosaury o třetinu delší, než je ten berlínský, dokládají existenci těch o třetinu těžších. Hmotnost narůstá s třetí mocninou délky. Máme li brachiosaura (nebo cokoli jiného) s délkou 1 a hmotností 1 a zvětšíme jeho délku na 1,13 (o 13%), hmotnost vzroste na 1,44 - tedy nejen o třetinu, ale téměř o polovinu větší.
Autorovou "nevinou" co se týče přehnaných odhadů si nelze být úplně jistý např. u délky udávané pro taxon Ceratosaurus ingens, známého pouze z velkých zubů. J. Mareš udává 17 metrů (a dokonce až 20 metrů u největších exemplářů) na základě nálezů britské expedice do Tendaguru v roce 1930, která zde měla objevit mimořádně velkou teropodí pánev a část páteře. Možná, že by tento extrémní odhad ospravedlnily sebrané spisy Fredericka Migeoda, vedoucího této expedice. Na druhou stranu, materiál C. ingens nikdy neobsahoval jiné fosilie než zuby a jejich fragmenty - pokud byly nalezená pánev a obratle skutečně teropodní, nikdy nebyly zařazeny pod C. ingens jako paratypy. Pochybnému ceratosaurovi (podle Rauhuta [1995] vykazují zuby spíše karcharodontosauridní znaky; stejně tak Holtz et al. [2004] taxon Megalosaurus ingens zahrnuje pod bazální tetanury) se věnuje také Paul (1988a), který jej sice uvádí mezi největšími dinosauřími predátory, uvádí však hmotnost pouze 5 tun (což určitě neukazuje na 17 m dlouhé zvíře) a zuby velikostně srovnává s daleko kratším tyrannosaurem. Bez znalosti polohy jednotlivých zubů v čelisti je dost dobře možné, že C. ingens byl ve skutečnosti menší, než devítimetrový C. nasicornis.
Podobně nepochopitelné jsou odhady pro 14 metrů dlouhého epanteria. Tuto délku zmiňuje pouze popisek k obrázku; v textu je rovnou zmiňován čistě spekulativní 15,5 m dlouhý rekordman - vzhledem k tomu, že už sám holotyp epanteria (zřejmě kolem 12 m dlouhý) je zřejmě pouze mimořádně velkým jedincem druhu Allosaurus fragilis, je takové svévolné nadsazení těžko obhajitelné. Kromě velikostních odhadů je navíc v kapitole o svrchnojurských predátorech Severní Ameriky komplikovaná i taxonomie. Obojí zrevidoval Mickey Mortimer v několika svých příspěvcích na DML (viz Zdroje) a v Theropod Database (viz postranní panel). J. Mareš vedle sebe uvádí druhy Allosaurus fragilis a Allosaurus atrox, přičemž taxon Epanterias amplexus považovuje za samostatný rod a druh a ztotožňuje s ním i "saurophaga", preokupované jméno pro nediagnostický holotyp allosaurida. Přestože se v této konkrétní části (str. 156 a 157) J. Mareš odvolává nejvíce na Bakkera, jde o téměř přesný přepis informací o těchto druzích z Paulova Predatory Dinosaurs of the World. Jediný významný rozdíl je v tom, že autor Záhady trvá na zachování epanteria jako samostatného rodu, zatímco Paul argumentoval pro svou kombinaci Allosaurus amplexus. I totožnost taxonu "Saurophagus" maximus s epanteriem je přejatá z Paula, ovšem informace o dvou neúplných kostrách už ne. Krom toho, že toto jméno (vztyčil Ray [1941]) je preokupované, jedná se o nomen nudum, ke kterému další autoři těžko mohli přiřazovat další materiál - nešlo by uspokojivě prokázat, že se nejedná o jakéhokoli jiného allosaurida. Až Hunt & Lucas (1987) ze saurophagova materiálu vztyčili lektotyp, ke kterému však už mezitím bylo přiřazeno značné množství dalšího materiálu. Mnoho z nich později Chure (1995) přiřadil ke svému nově zavedenému taxonu Saurophaganax maximus, jehož holotyp - část dorzálního obratle - opět pocházela z materiálu přiřazenému k "saurofágovi". Přesto se nejednalo o jeho náhradu, protože lektotyp není diagnostický a nedovoluje tedy oba taxony synonymizovat. Ve stejné práci navíc Chure ukázal, že ačkoli Saurophaganax (a tedy také přinejmenším část materiálu "saurofága") disponuje autapomorfiemi, které ho vymezují proti A. fragilis, Epanterias naopak unikátní znaky postrádá. Tato argumentace odporuje Bakkerovým argumentům o výskytu, ale drží se jí i jiní autoři, zahrnující epanteria pod druh A. fragilis (Holtz et al. 2004). Smith (1996, 1999) rovněž podložil tvrzení, podle kterého nelze rozdíly pozorované mezi A. fragilis a A. atrox považovat za dostatečné k druhové separaci.
Epanterias amplexus tak může být synonymem kteréhokoli z jiných, autorem rozlišovaných morrisonských allosauridů, nebo, ze současného pohledu, S. maximus a A. fragilis. Neexistuje evidence pro 15 (nebo, když jsme u toho, 14) metrů dlouhého jedince - Mortimerův odhad odvozený z korakoidu zní 12,1 metru. Materiál "saurophaga" historicky zahrnoval směs z různých bazálních tetanur, snad včetně karcharodontosaurida a spinosaurida, a zřejmě daleko více než jen 2 kostry. Materiál přiřazený k saurophaganaxovi, konkrétně humerus - před Chureovou revizí zřejmě rovněž tvořící součást "saurophaga" - skutečně ukazuje až na délku 13 m, Mortimerovy odhady postavené na femuru se však pohybují mezi 10,2 - 10,9 metry.
Dalším teropodem, který to, co se týče velikostních odhadů, "odnesl", je Deinocheirus mirificus. Je zřejmé, že u taxonu, jehož jediný známý materiál tvoří s výjimkou několika dalších fragmentů pouze dva obrovské skapulakorakoidy, závisí odhady délky ještě silněji na systematice a fylogenetické pozici. J. Mareš deinocheira porovnává s velociraptorem a allosaurem, a vyvozuje, že zatímco v prvním případě bychom měli 14 m dlouhého dromaeosaurida, pokud by se D. mirificus podobal více tomu druhému, mohl by měřit okolo 20 metrů. Kde je podklad pro tyto možnosti zůstává záhadou. Na G. S. Paula se to tentokrát svést nedá - ten pouze uvádí, že někteří nejmenovaní autoři došli k fantastickým rozměrům extrapolací deinocheirových rozměrů z poměru vůči předním končetinám tyrannosaura (Co k tomu tyto autory asi vedlo? Zatímco přední končetiny deinocheira jsou zjevně tříprsté, pro tyrannosauridy je typická funkční didaktylie - nemluvě o dalších apomorfiích tyrannosauridů, jako je celkově redukovaný karpus). Pochybuji také, že materiál deinocheira vykazuje nějakou zvláštní podobnost s allosauridy. J. Mareš zmiňuje možnost ornitomimidní (nebo, přesněji, ornitomimosauří) příslušnosti deinocheira, ale zamítá ji - spolu s Czerkasovými idejemi o býložravosti - ve prospěch větší podobnosti s dromaeosauridy, především na základě drápů. Je pravda, že drápy jsou nejsilnější evidencí pro deinocheirovu predaci a že postrádají pro ornitomimosaury typický distálně umístěný hrbolek ohybače (Makovicky et al. 2004), přesto se však právě taková deinocheirova fylogenetická příslušnost zdá být podpořena největším počtem důkazů (mezi které patří např. redukovaný deltopektorální hřeben nebo téměř shodná délka metakarpálů I, II a III), jak si povšiml větší počet autorů (Ostrom 1972; Paul 1988a; Barsbold & Osmólska 1990 - už v původním popisu [Osmólska & Roniewicz 1970] je nicméně zaznamenáno, že "the structure of the manus and humerus strongly resembles that of Ornithomimus"). Povrdily ji i kladistické analýzy (Pisani et al. 2002), přesto však detaily o ekologii deinocheira zůstávají tajemstvím - v tom lze s J. Marešem souhlasit i po více než patnácti letech. Pokud jde o velikost, ta se zřejmě pohybovala pod 14 metry - zde ale záleží na tom, který ornitomimosauří druh použijeme pro extrapolaci rozměrů.
Jako poslední případ velikostně nadhodnocených teropodů je možno uvést vždy populárního tyrannosaura. J. Mareš např. uvádí nález 1,75 m dlouhé lebky v Montaně roku 1987, která má být součástí 15 m dlouhé kostry. Problém začíná už v samotném faktu, že ve zmiňovaném roce v Montaně žádná kostra nalezena nebyla - alespoň podle katalogu dostupného tyrannosaurova materiálu. O rok později byl sice v Montaně skutečně objeven exemplář, sestávající z relativně kompletní kostry (MOR 555, "the Wankel rex" - zdá se, že jde skutečně o exemplář odkazovaný v Záhadě, protože právě tento materiál potvrdil redukovaný počet prstů na tyrannosaurově přední končetině), ale přestože rozměry jeho lebky není snadné dohledat, kostra jako taková určitě neměří 15 m: femur má délku 1280 mm, zatímco u Sue - největšího známého exempláře tyrannosaura, dlouhého 12,8 metru - femury měří 1321 (pravý) a 1308 (levý) mm; komparací z tohoto údaje dojdeme k délce asi 12,45 metru. 1,75 metru dlouhá lebka tyrannosaura má blíže spíše ke sci-fi než realitě; vyrovnala by se délklu daleko méně masivní lebce spinosaura, největšího známého teropoda vůbec. Lebka Sue měří 141 cm a nejdelší známá tyrannosaurova lebka vůbec, MOR 008, 150 cm (už tento údaj dává prostou extrapolací hodnotu 13,6 m). Mortimer udává jako délku lebky Sue 139,4 cm, což by odhad dále zvýšilo na 13,8 metru. Zároveň však připomíná, že jednotlivé tyrannosauří exempláře nemají stejné proporce - pokud budeme porovnávat jednotlivé části těla, došly bychom z nich k odlišným poměrům, a žádný z nich nakonec nemusí vypovídat o celkové délce těla. MOR 008 má např. maxillu jen z 84% tak dlouhou, jako je ta u Sue, a zubní řada zabírá délkou 90% z té u Sue. Ani tato lebka tak nemusela patřit exempláři delšímu než 12,8 metru. Existuje však evidence pro daleko větší lebky, logicky se ale objevuje pochybnost, zda by i ony šly ještě přiřadit druhu T. rex - přestože infraspecifická velikostní variace nemůže být zanedbávána. Existuje izolovaná ozubená maxilla s označením UCMP 118742, dlouhá 810 mm. Paul (1988a) stanovil, že jedinec tohoto druhu mohl mít skutečně 175 cm dlouhou lebku, s celkovou délkou těla kolem 13,6 metru, hmotností 12 tun a délkou femuru získanou extrapolací z holotypu asi 1675 mm. Ve stejné práci autor dokonce vyvodil závěr, že pokud tento materiál skutečně patří druhu T. rex, pak všichni ostatní známí jedinci musí být subadultní. Průměrná hmotnost dospělého tyrannosaura by se pak posunula k 15 tunám, s tím, že rekordní jedinci by dosahovali 20 tun. Všechny tyto údaje se však zdají být extrémně nadhodnocené - ve srovnání s holotypem tyrannosaura AMNH 5027 (maxilla dlouhá 710 mm) není tato fosilie větší o 29%, jak udává Paul, ale pouze o 14%. Ve svém příspěvku na DML z roku 1996 Paul uznal, že původní hmotnostní odhad byl nadhodnocený, a Mortimerova databáze uvádí podstatně realističtější odhad pro délku lebky, do které tato maxilla patřila - asi 131 cm, což by ukazovalo na délku kolem 12,1 - 12,4 metru - fakticky menší, než u Sue. Ani další tyrannosauří exempláře v knize však na tom nejsou o nic lépe. Např. "Black Beauty" (RTMP 81.6.1), uložená v Tyrellově muzeu (z knihy to není zcela zřejmé, ale jedná se o totožnou kostru, jako na straně 211), má být 14 m dlouhá - ve skutečnosti měří 11,7 metru a je výrazně kratší než Sue. Ta mimochodem také nezůstává v knize ušetřena - přezdívka exempláře FMNH PR2081 zde sice není zmíněna, z příběhu o právních sporech a zmínce o Sue Hendrickson je ovšem celá záležitost naprosto jasná. Tento etalon zdejšího poměřování tyrannosaurů - výborně zachovalá a zároveň největší kostra - je naprosto nezpochybnitelně dlouhý 12,8 metru, v knize se ovšem vyskytuje hodnota neuvěřitelných 16 metrů, kterou můžeme přirovnat snad pouze ke spinosaurovi. Kde se tento odhad vzal? Těžko říct; Mortimer ale připomíná proměnlivé proporce tyrannosauridů a uvádí, že kdybychom extrapolovaly rozměry Sue podle velikosti maxilly z holotypu (rozdíl 710 vůči 861 milimetrům), došli bychom k délce asi 15,5 metru. Chyba spočívá v tom, že prakticky libovolná jiná část kostry dá naprosto odlišné výsledky - druhý metatarzál je dokonce u Sue o 5% kratší než u holotypu! A ani objevy, učiněné až po sepsání záhady, nevypovídají o tyrannosaurech delších, než je Sue. "Celeste" neboli "C-rex" (MOR 1126) měla být podle slov J. Hornera o 10% delší než Sue (asi 14,1 metru), problémem jsou však opět proměnlivé proporce - několik kostí spodní čelisti, části páteře, lopatka, pánev, sakrum a dvacítka žeber se zdá být příliš málo na odvození spolehlivých údajů. Nové Mortimerovy údaje udávají 12,3 metru.
Jednoduše řečeno, evidence pro 13+ metrového tyrannosaura naprosto chybí. Pokud jde o roztržení tyrannosaura na taxony Tyrannosaurus rex a Dynamosaurus imperiosus: objektivní kritérium pro rozlišení druhů ve fosilním záznamu bude samozřejmě vždycky chybět, protože nevíme, mezi kterými populacemi už existovala reprodukční bariéra, kde se jedná o polymorfizmus, jaké typy speciačních událostí probíhaly apod. Zbývá několik způsobů, jak rozeznat fosilní druh - jeden z nich je založen na čase, konkrétně době, po kterou existuje jedna populace. Taxon, který je nalézán v sedimentech s dlouhou depoziční periodou (miliony let), zřejmě nereprezentuje jeden druh v tom smyslu, jak tento pojem chápe biologie zkoumající živé organismy. Dalším měřítkem je hledání odlišujících znaků, autapomorfií. A zde se objevuje problém - je zřejmé, že některým autorům bude pro uznání nového druhu stačit méně apomorfií než jiným. Na tuto otázku by se dal napsat článek téměř stejně obsáhlý jako tento, vysvětlující, proč jsou ne-monotypické rody nevýhodné (implikují totiž fylogenetickou hypotézu samotným pojmenováním taxonu), a proč je dobré pojmenovat druh, i když je založen na extrémně fragmentárním materiálu, pokud disponuje přesvědčivými autapomorfiemi. Autorům, kterým stačí pro rozeznání nového druhu menší množství apomorfií, se říká "splitteři" - je logické, že paleontologové s tímto přístupem budou rozeznávat více nezávislých druhů, než jejich kolegové; a v jistém smyslu tak budou "roztrhávat" existující taxony. (Co se týče rodového jména, to už je víceméně formalita - neexistuje žádné kritérium, které by říkalo, kolik rozdílů je nutných pro specifickou a kolik pro generickou separaci, takže pokud máme druhy D. imperiosus a T. rex, faktický rozdíl oproti T. imperiosus a T. rex je nepatrný). Bez možnosti studovat ontogenezi a populační biologii daného organismu navíc nezjistíme, které autapomorfie vymezují populace, a které pouze pohlaví nebo ontogenetická stadia. Dale Russell, "splitter", zastává názor, že rozdíly mezi "dynamosaurem" a "klasickým" tyrannosaurem jsou dostatečné pro validitu obou taxonů. Většina paleontologů v době vydání Záhady se však domnívala, že tyto rozdíly lze vysvětlit sexuálním dimorfizmem, alometrií (= změnami v celkových proporcích během ontogeneze) nebo individuálními variacemi, a dnes už opozice k tomuto názoru ani neexistuje. Jméno Dynamosaurus imperiosus tak vymizelo z literatury zřejmě definitivně.
Ilustrace
Za ilustracemi v knize stojí Barbora Kyšková, malířka a autorka ilustrací pro větší množství knih z různých oblastí. Umělecký dojem je samozřejmě čistě subjektivní záležitost. Mně se malby i kresby poměrně líbí, nemám např. důvod B. Kyškové něco vytýkat ve srovnání se Zdeňkem Burianem. Pozadí samozřejmě není tolik propracované, ale může se jednat stejně dobře o nevýhodu jako výhodu - ilustrace tak dává více vyniknout samotnému zvířeti. Osobně si myslím, že oba malíři ani omylem nedosahují kvality současných autorů fotorealistických maleb, využívajících počítačové grafiky, jako je Raúl Martín nebo Luis Rey. Skutečně nevím, jaký jiný dojem bych měl posuzovat, než to, nakolik ve mně obraz dokáže vzbudit zdání skutečnosti.
To, co už samozřejmě objektivně posoudit lze, je, nakolik ilustrace odpovídají realitě, a v jistém smyslu i jejich originalita. Tady by B. Kyškové příslušelo podstatně nižší hodnocení. Mezi ilustrace, které jsou podle mého názoru skutečně pěkné a zároveň jim není mnoho co vytknout, patří např. perokresba malých teropodů ze strany 121 (dobrá, chybí peří, ale tady je to spekulativní), Stegosaurus "ungulatus" na straně 149, Parasaurolophus walkeri ze strany 176, Styracosaurus albertensis ze strany 187 nebo většina perokreseb ceratopsích lebek a sauropodů. Mnoho dalších je ale anatomicky přinejmenším nepřesných. Mezi ně patří většina ilustrací teropodů, jejichž přední končetiny měly směrovat dlaněmi spíše dovnitř než dolů. Některé neproporcionální tělesné části nebo nepřesná textura kůže se zřejmě nedají popřít ani u jiných ilustrací. Plateosaurus (jako bazální sauropodomorf) a Segnosaurus (jako maniraptor) měli být bipední, ne kvadrupední. Styracosaurus ze strany 187 má příliš malou pre-skvamosální část lebky. Některé další ilustrace jsou zpochybnitelné z hlediska chování "účastníků" - to platí pro bojující barosaury ze strany 132, diplodoky spásající koruny stromů ze strany 134 a lebkami do sebe narážející pachycephalosaury ze stran 180 a 181 (těmi se zabývaly i seriózní biomechanické studie, aby došly k závěru technické nemožnosti takového chování). Mimořádně nepovedení a až odpudiví jsou deinonychové ze strany 164, kteří vypadají jako nepovedené zmenšeniny tyrannosaurů a vrásčitá šedomodrá kůže (na malbách B. Kyškové poměrně oblíbená) jim také příliš nepřidává. Lebka má zcela špatný tvar a podivné se zdá být i skloubení rukou. Totéž, byť ne tak silně, platí i o hypotetickém dromaeosauridním deinocheirovi ze strany 218. Malba "dynamosaura" na straně 237 má zcela chybný tvar lebky. Premaxillární část je moc vysoká a lebka díky tomu působí dojmem, jako by od spánku ke špičce čelisti vůbec neklesala, což neodpovídá realitě (jak lze snadno zjistit prohlédnutím si dobře zachovalé lebky. Profil dolní čelisti je úplně špatně - kost zubní se nezvyšovala schodovitě směrem ke skloubení čelisti, ale byla po celé délce prohnutá dolů. Naopak v místě kontaktu kosti zubní s angularem a surangularem se tloušťka spodní čelisti prudce zvyšuje, a šikmo vzato (po linii kontaktu) se zde dokonce vyrovnává horní čelisti. K místo skloubení se spodní hrana dolní čelisti zvedá více prudce a zároveň po větší délce, než rekonstrukce ukazuje. Spodní čelist nanotyranna na straně 228 by na tom byla lépe, kdyby měla patřit dospělému tyrannosaurovi - naneštěstí tomu tak není a hlavě se dá opět spousta věcí vytknout. Měla být více protáhlejší, s celkově daleko štíhlejším čenichem a menší vzdálenosti od horního okraje očnice ke spodní hraně horní čelisti. I spodní čelist měla být daleko protáhlejší a její zesílení nemělo být patrné pouze pod očnicemi, jak ukazuje ilustrace, ale po celé délce od místa pod spánkem až ke kontaktu mezi maxillou a jugalem. Spinosaurovi ze strany 173 nelze vyčítat absenci typického krokodýlího rostra, protože lebeční materiál této bazální tetanury nebyl dostatečně kompletní. Hlava na této konkrétní ilustraci ovšem není postavena vůbec na žádném základě, včetně staré Stromerovy rekonstrukce, a ani příliš nepřipomíná teropody.
To, co už samozřejmě objektivně posoudit lze, je, nakolik ilustrace odpovídají realitě, a v jistém smyslu i jejich originalita. Tady by B. Kyškové příslušelo podstatně nižší hodnocení. Mezi ilustrace, které jsou podle mého názoru skutečně pěkné a zároveň jim není mnoho co vytknout, patří např. perokresba malých teropodů ze strany 121 (dobrá, chybí peří, ale tady je to spekulativní), Stegosaurus "ungulatus" na straně 149, Parasaurolophus walkeri ze strany 176, Styracosaurus albertensis ze strany 187 nebo většina perokreseb ceratopsích lebek a sauropodů. Mnoho dalších je ale anatomicky přinejmenším nepřesných. Mezi ně patří většina ilustrací teropodů, jejichž přední končetiny měly směrovat dlaněmi spíše dovnitř než dolů. Některé neproporcionální tělesné části nebo nepřesná textura kůže se zřejmě nedají popřít ani u jiných ilustrací. Plateosaurus (jako bazální sauropodomorf) a Segnosaurus (jako maniraptor) měli být bipední, ne kvadrupední. Styracosaurus ze strany 187 má příliš malou pre-skvamosální část lebky. Některé další ilustrace jsou zpochybnitelné z hlediska chování "účastníků" - to platí pro bojující barosaury ze strany 132, diplodoky spásající koruny stromů ze strany 134 a lebkami do sebe narážející pachycephalosaury ze stran 180 a 181 (těmi se zabývaly i seriózní biomechanické studie, aby došly k závěru technické nemožnosti takového chování). Mimořádně nepovedení a až odpudiví jsou deinonychové ze strany 164, kteří vypadají jako nepovedené zmenšeniny tyrannosaurů a vrásčitá šedomodrá kůže (na malbách B. Kyškové poměrně oblíbená) jim také příliš nepřidává. Lebka má zcela špatný tvar a podivné se zdá být i skloubení rukou. Totéž, byť ne tak silně, platí i o hypotetickém dromaeosauridním deinocheirovi ze strany 218. Malba "dynamosaura" na straně 237 má zcela chybný tvar lebky. Premaxillární část je moc vysoká a lebka díky tomu působí dojmem, jako by od spánku ke špičce čelisti vůbec neklesala, což neodpovídá realitě (jak lze snadno zjistit prohlédnutím si dobře zachovalé lebky. Profil dolní čelisti je úplně špatně - kost zubní se nezvyšovala schodovitě směrem ke skloubení čelisti, ale byla po celé délce prohnutá dolů. Naopak v místě kontaktu kosti zubní s angularem a surangularem se tloušťka spodní čelisti prudce zvyšuje, a šikmo vzato (po linii kontaktu) se zde dokonce vyrovnává horní čelisti. K místo skloubení se spodní hrana dolní čelisti zvedá více prudce a zároveň po větší délce, než rekonstrukce ukazuje. Spodní čelist nanotyranna na straně 228 by na tom byla lépe, kdyby měla patřit dospělému tyrannosaurovi - naneštěstí tomu tak není a hlavě se dá opět spousta věcí vytknout. Měla být více protáhlejší, s celkově daleko štíhlejším čenichem a menší vzdálenosti od horního okraje očnice ke spodní hraně horní čelisti. I spodní čelist měla být daleko protáhlejší a její zesílení nemělo být patrné pouze pod očnicemi, jak ukazuje ilustrace, ale po celé délce od místa pod spánkem až ke kontaktu mezi maxillou a jugalem. Spinosaurovi ze strany 173 nelze vyčítat absenci typického krokodýlího rostra, protože lebeční materiál této bazální tetanury nebyl dostatečně kompletní. Hlava na této konkrétní ilustraci ovšem není postavena vůbec na žádném základě, včetně staré Stromerovy rekonstrukce, a ani příliš nepřipomíná teropody.
Pokud jde o snahu být originální a představit dinosaury "v novém světle" i vizuálními prostředky, ta je samozřejmě nejsnadnějším terčem kritiky. Chmýří na krku a hřbetu ptakopánvých (strana 253) a bazálních dinosauromorfů (strana 103) by šlo kritizovat jako zcela nepodložené ještě před rokem, situace se však obrátila a po objevu tianyulonga se něco takového zdá nejen možné, ale i pravděpodobné. Tito a mnoho dalších částečně opeřených dinosaurů je silně inspirováno prací G. S. Paula. Problém je v tom, že Paul se rozhodl, že bude malovat s peřím všechny dravé dinosaury, u kterých není důkaz o opaku (v jeho knize tedy nenajdeme opeřené tyrannosauroidy ani abelisauridy). J. Mareš se však zřejmě rozhodl nabídnout ve své knize vyrovnanější pohled, a tak některé ilustrace jsou provedené s peřím, jiné ne. Výběr je ale často chybný. Jistě, autor ani ilustrátorka nemohli vědět, jaké peří bude nalezeno u kterých teropodích kladů, ale mohli dojít k předpokladu - implicitně obsaženém v Paulově knize - že největší vyspělost (nebo pravděpodobnost přítomnosti) peří se dá předpokládat u linií fylogeneticky bližších k ptákům. Zatímco Paulovy kresby byly ohledně peří konzistentní (tj. tyrannosauroidi jej neměli, ornithomimidi ano), Záhada není - Gallimimus bullatus na straně 217 peří na krku a zádech má, struthiomimové na perokresbě ze strany 251 a Dromiceiomimus ze strany 266 ne. Jistě, v současnosti víme, že přítomnost peří byla proměnlivá i mezi zástupci jednoho poměrně úzkého kladu (Compsognathidae, např.), ale ilustrace v Záhadě tímto ztrácejí na propracovanosti. Když se nad obrázky hlouběji zamyslíme, zjistíme, že podle Záhady se charakteristika tělesného pokryvu nezměnila po 170 milionů let, od střednětriasového lagosucha, který měl s ptáky společného jen málo, až po svrchnokřídového deinonycha, kterého bychom už klidně mohli ptákem nazývat (viz strana 255) - oba disponují pouze jakýmsi ochmýřením vzadu za hlavou. Paul si v Predatory Dinosaurs of the World dovolil s peřím experimentovat: jeho velociraptorové jsou celí pokryti vláknitým protopeřím, s chocholkou obrysového peří vzadu na hlavě (strana 172), Ornitholestes je rovněž pokrytý peřím od hlavy až po ocas (strana 211), a Coelophysis má za hlavou výraznou pernatou chocholku, kombinovanou s jakousi hřívou v místech, kde se krk napojuje na tělo (strana 264). Pokud z opeřených dinosaurů v Záhadě máme rozporuplné dojmy, chobot giraffatitana (strana 60) nebo "ultrasaura" (strana 137) lze v klidu odmítnout. Darren Naish napsal o sauropodích chobotech na svém blogu vynikající článek, kde tuto možnost kompletně vyvrací. Není zde zapotřebí přepisovat všechny jeho argumenty, uvedu tedy jen, že seriózní výzkum kraniální neurologie neprokázal přítomnost hypertrofovaných nervů, nutných k inervaci chobotu u současných živočichů. Sauropodi s choboty jsou tak jednou z těch spekulativních myšlenek, jejich zahrnutí do Záhady se při pohledu z dnešní doby nevyplatilo. Vertikálně vztyčené krky brachiosauridů opět patří do kategorie spekulací, které by šlo kritizovat ještě před rokem: práce Taylor et al. 2009 sice není nijak definitivní, prokazuje ovšem, že paleontologická komunita tuto možnost nemůže zavrhnout.
Co ovšem ilustrace v knize sráží skutečně velkou měrou je fakt, že mnoho z nich je doslova vykradeno z jiných prací, aniž by byli původní autoři jakkoli kreditováni. Platí to naneštěstí zrovna pro ty obrázky, které budou čtenářům připadat nejpovedenější. Následující seznam nemá být a ani není vyčerpávající, ukazuje jen příklady ilustrací, okopírovaných od jiných autorů. Perokresba gasosaura ze strany 130 vykrádá dílo Johna Sibbicka, jen je vypuštěno pozadí. Paulova kresba ornithomimů (strana 134) má v Záhadě změněné pozadí a velmi lehce modifikovanou polohu druhého jedince, aby byla zrcadlově obrácena a prohlášena za ilustraci gallimimů - jediný faktický rozdíl spočívá v trochu jiném úhlu stehna svíraného s tělem. Téměř přesně je vykradena kompozice ilustrace zobrazující útok allosaurů na A. ajax (strana 143 v Záhadě), a to z Paulovy kresby allosaurů útočících na druh A. louisae (strana 45). Drobné detaily (poloha končetin apatosaura, poloha útočících allosaurů) se liší. Anchiceratops ze strany 197 je opět okopírován od Sibbicka. Perokresba ze strany 212 je okopírována s výjimkou velice drobných detailů od G. S. Paula (1988a:20). Perokresba ze strany 212 je věrnou kopií té Paulovy ze strany 301. I malba albertosaura na protější straně je vykradena od Paula (1988a:330) s tou výjimkou, že byla zrcadlově obrácena. Stejný "trik" byl použit i na hlavu oviraptora ze strany 221 (v Paulovi 376). Kolik ilustrací je převzato z Bakkerovy knihy The Dinosaur Heresies nemůžu říct, protože ji nemám k dispozici.
Ke knize
Dosud jsem se zabýval uváděním na pravou míru některých omylů nebo nepřesných a zastaralých údajů. Uvědomuji si, že tato část mohla znít v mnohém kriticky k autorovi, a někdy se skutečně o kritiku jednalo - ve většině případů však ne. Dinosauří paleontologie prošla v uplynulých 17 letech velkými změnami a to, co dnes považujeme za fakty, bude v mnoha případech během následujících 17 let také vyvráceno. Uvidíme syntézu paleontologie s evolučně developmentární biologií, molekulární biologií, genomikou a proteomikou? Růst výpočetního výkonu a rostoucí dostupnost špičkových zobrazovacích technologií, jako jsou CT skeny s vysokým rozlišením, také mohou paleontologii pouze pomoci. Nemohu také neuvést, že je snadné kritizovat Záhadu z pozice někoho, kdo může získat téměř libovolnou informaci z PDF uložených na disku a trochy snahy při zacházení s Googlem. Jaroslav Mareš často čerpal své informace z non-komformních zdrojů, což se z dnešní pozice někdy vyplatilo, někdy ne. Těžko mu může kdokoli vyčítat, že hypotézy o příbuzenství ve své knize předložil na základě Paula a Bakkera místo Gauthiera. Na druhou stranu je možné říct, že Bakkerův kladogram na poslední stránce mohl být vypuštěn; autor konec konců připustil, že se nechce zabývat systematickými záležitostmi, protože na ty se názory liší autor od autora. Dnes už konkrétně tento Bakkerův stromek napáchá více škody než užitku: např. návštěvnící Zoo Dvůr Králové, kde je vyvěšena jeho nástěnná verze, jsou s překonanými fylogenetickými hypotézami (Phytodinosauria, Orbitoscuta) konfrontování zcela zbytečně.
Pokud jde o kvality knihy jako takové, čtivost patří na přední příčku. Devadesát procent toho, co o dinosaurech (ať už jako celku, nebo o jednotlivých druzích) víme, se týká detailní anatomie a fylogeneze, autor však dokázal napsat obsáhlou knihu bez toho, aby se oběma kategoriím musel věnovat v nějaké zvláštní šířce. Čtenář se sice dozví, v čem konkrétně spočívá rozdíl mezi "plazím" a "ptačím" typem pánve, a jaké názory na vývoj a vzájemná příbuzenství dinosaurů mají (měli) Paul a Bakker, většinu obsahu knihy však tvoří atraktivnější informace - o ekologii nebo o časovém a geografickém rozšíření (kteří býložravci byli kořistí kterých dravců, které druhy byly natolik úspěšné, že jejich zástupce nalézáme na více kontinentech, apod.). Jak dinosauři vznikli a vydobyli si vedoucí místo na souších, jak svou pozici upevnili v křídě a jak dosáhli úžasného a v evoluci suchozemských obratlovců dosud nevídaného rozkvětu v křídě - to jsou dost možná nejkvalitnější části knihy. Příběhy velkých paleontologů, Mantellem a Bucklandem počínaje, Copem a Marshem pokračujíce a Nopcsou, Sternbergem, Huenem, Osbornem a Brownem konče - s výjimkou drobnějších příběhů Bakkera, Hornera, Gilletteho, Currieho, Serena, Donga a mnoha dalších - jsou trochu spornější záležitostí. Čtivost, zábavnost a zajímavost se jim nedá upřít - nedivil bych se, kdyby u mnoha čtěnářů byly příběhy příležitostného balkánského špióna Franze von Nopcsy nebo znepřátelených rivalů Marshe a Copea nejoblíbenější částí knihy. Ale podobné vyprávění je už vždy více o lidech než dinosaurech. Chápu, že mnoho lidí by nesouhlasilo a obhajovalo názor, že není možné psát o paleontologii bez zmínky o paleontolozích, a nakonec bych s nimi i souhlasil. Jediné, co bych přidal, je příběh prvního návrhu na dinosauří původ ptáků, související neodmyslitelně s takovými jmény, jako jsou Gegenbaur, Huxley, Haeckel, Cope a Marsh. Prvkem autorské licence jsou (rádoby) poetické popisy krajiny, ve které dinosauři žili, a příběhy jednotlivých dinosaurů, zpravidla konfrontace dravců a jejich kořisti. Nemyslím si, že by tyto části knize přinášely výraznou přidanou hodnotu, asi ale dovedou určitý typ čtenářů oslovit a v každém případě jí nijak neškodí.
To už nedokáži s takovou jistotou říct o kapitolách "Mohli někde přežít dodnes?" a "Dino sapiens". První z nich se zabývá kryptozoologickými spekulacemi, druhá možnou evolucí dinosaurů v inteligentní formy připomínající Homo sapiens. Osobně bych v jejich hodnocení byl velice opatrný. Ten, kdo zná dílo Jaroslava Mareše trochu podrobněji, ví, že kryptozoologie patří do jeho aktivních zájmů přinejmenším stejně jistě jako paleontologie. "Aktivní zájem" je zde možná pořád trochu slabý výraz, neboť J. Mareš byl dokonce organizátorem několika expedic pátrajících po "kryptidech", tedy zvířatech, jejichž existence není podložena vědecky dostatečnou evidencí. Já osobně jsem shledával kapitolu o možném přežití non-avianních dinosaurů stejně zajímavou jako jiné části knihy a protože (nebo ačkoli?) jde o populárně naučnou publikaci, nemyslím si, že je její zahrnutí vyloženě nevhodné. Že v pralesech střední Afriky a Jižní Ameriky - v obou případech bývalé Gondwany - existují ekosystémy v mnoha směrech nedotčené katastrofou na přelomu křídy a paleocénu, je nezpochybnitelný fakt. Jsem přesvědčený, že i přes lákavou a solidně znějící podporu, kterou J. Mareš hypotéze o přežití nepatčích dinosaurů v tamních končinách přináší, se racionálně uvažující člověk musí zamyslet nad pravděpodobností hypotézy, že by zde izolované skupinky dinosaurů přežili všechny změny posledních 65 milionů let, včetně ekologického rozmachu savců, a odmítnout ji. Ačkoli nevěřím v hypotézu, kterou J. Mareš předkládá, jde o velice lákavou a zábavnou myšlenku, kterou bych nikomu nezakazoval se zabývat, a to ani v naučné publikaci, jakou Záhada dinosaurů je. Bylo by levné snažit se kritizovat knihu takové úrovně, kterou Záhada je, z pozice člověka znechuceného nad tím, že "autor je kryptozoolog a tedy pavědec".
Ani kapitola "Dino sapiens", zamyslíme-li se nad ní hlouběji, není nijak zvlášť nevhodná pro populárně-naučnou publikaci. Jistě, jde víceméně o fantazie s nulovou podporou fosilního záznamu, jejichž příčina je pouze v lidské fascinaci dinosaury a snaze projektovat do nich vlastnosti dnešního světa, v tomto případě lidskou inteligenci. Ale zabývali se jí seriózní vědci, včetně Dalea Russella, nebo - samozřejmě dlouho po napsání Záhady - Darrena Naishe (na svém blogu). Je ale nutné připomenout, že pro oba se jednalo o jakýsi zábavný "vedlejšák", ne předmět seriózního výzkumu, a tak by měl být brán. Russellův slizoid je dnes naprosto neaktuální; někteří by však jistě byli toho názoru, že působivost mu nejde upřít. Víme, že tělo troodontidů bylo pokryto obrysovým peřím, víme (a tohle mohl vědět i Russell), že pro zvětšení mozkovny není nijak nutná vertikalizace páteře a ztráta ocasu - tyto tendence dávají smysl u primátů, ne ale u maniraptorů. Ovšem jedna skutečně důležitá hranice, která v žádném případě neměla být překročena, je ta mezi inteligentními troodontidy a šediváckými "ufouny". Jde přesně o tu hranici, za kterou si již pravděpodobně většina čtenářů začne klepat na čelo, a podle mého názoru naprosto oprávněně. Populárně-naučná publikace s několika zábavnými spekulacemi, dobrá. Populárně-naučná publikace kombinovaná samoúčelně se špatnou sci-fi povídkou, ne! Z textu lze usoudit, že J. Mareš má nebo měl s "ufologií" alespoň okrajově cosi do činění ("Podotýkám jen pro úplnost, že [podobnost mezi ufouny a inteligentním dinosauroidem] nebyla dosud nikde publikována a že jsem si jí povšiml náhodně při studiu od problematiky dinosaurů zcela odlišného materiálu." - str. 271). Pokud ano, nelíbí se mi, že ji zamíchal do knihy o dinosaurech. Kryptozoologie není věda; nepracuje vědeckými metodami a nežádá evidenci dostatečnou pro vědu, může však být užitečná při hledání nových druhů, obzvláště, když ji dělá zoolog (jako byl Bernard Heuvelmans). Ale to, čím se J. Mareš v Záhadě zabývá na stranách 269 - 271, je o mnoho stupňů horší.
Vidím, že jsem si nejsilnější kritiku nechal na konec, což má logicky za následek, že recenze bude působit, jako by knihu odmítala. Ve skutečnosti jsem přesvědčen, že klady u Záhady daleko převyšují všechny zápory, co se mi podařilo najít: kniha je zajímavá a až na ty nejvíce spekulativní části poměrně věrně reflektuje stav dinosauří paleontologie ve své době (je ovšem nutno poznamenat - už jen pro sílu inspirace, kterou tito Záhadě byli - že v lecčems reflektuje spíše názory Bakkera a Paula, které často v souladu s většinou dalších paleontologů nebyly). V současnosti je však mnoho jejích informací zkreslených a určitě by si zasloužila nástupce, který by ukázal, jak dinosauří paleontologie vypadá koncem první dekády 21. století. Kdyby za tímto nástupcem stál Jaroslav Mareš, o to lépe.
Zdroje:
- http://dml.cmnh.org/2003Jul/msg00355.html
- http://dml.cmnh.org/2001Jan/msg00445.html
- Allain, R. & Aquesbi, N. 2008. Anatomy and phylogenetic relationships of Tazoudasaurus naimi (Dinosauria, Sauropoda) from the late Early Jurassic of Morocco. Geodiversitas 30(2):345-424.
- Arcucci, A. B. 1998. New information about dinosaur precursors from the Triassic Los Chanares Fauna, La Rioja, Argentina. In: Journal of African Earth Sciences, v. 27, n. 1A July 1998, Special Abstracts Issue, Gondwana 10: Event Stratigraphy of Gondwana: 9-10.
- Bakker, R.T. 1986. The Dinosaur Heresies: New theories unlocking the mystery of the dinosaurs and their extinction. New York: William Morrow & Co, 481 p.
- Barrett, P. M. 2009. The affinities of the enigmatic dinosaur Eshanosaurus deguchiianus from the Early Jurassic of Yunnan Province, People's Republic of China. Palaeontology 52 (4):681-688.
- Barsbold, R. 1983b. On some avian features in the morphology of theropods. Transactions of the Joint Soviet–Mongolian Paleontological Expedition. 24:96–103 (in Russian).
- Barsbold, R. & Osmólska, H. 1990. Ornithomimosauria. In D.B. Weishampel, P. Dodson & H. Osmólska (eds.), The Dinosauria, 225–244. University of California Press, Berkeley: 225-244.
- Benton, M. J. 1999. Scleromochlus taylori and the origin of dinosaurs and pterosaurs. Philosophical Transactions of the Royal Society of London B. 354:1423-1446.
- Bininda-Emonds, O. R. P, Cardillo, M., Jones, K. E., MacPhee, R. D. E., Beck, R. M. D., Grenyer, R., Price, S. A., Vos, R. A., Gittleman, J. L. & Purvis, A. 2007. The delayed rise of present-day mammals. Nature 446:507-511.
- Bonaparte, J. F., Novas, F. E. & Coria, R. A. 1990. Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia. Contributions in Science Natural History Museum Los Angeles County, 416:1-42.
- Calvo, J. O. & Salgado, L. 1995. Rebbachisaurus tessonei sp. nov. A new Sauropoda from the Albian-Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. Gaia 11: 13–33.
- Cooper, M. R. 1985. A revision of the ornithischian dinosaur Kangnasaurus coetzeei Haughton, with a classification of the Ornithischia. Annals of the South African Museum 95 (8):281-317.
- Galton, P. M. 1985. The poposaurid thecodontian Teratosaurus suevicus v. Meyer, plus referred specimens mostly based on prosauropod dinosaurs, from the Middle Stubensandstein (Upper Triassic) of Nordwurttenburg. Stuttgarter Beitrage zur Naturkunde (ser. B) 116:1-29.
- Gauthier, J. A. 1984. A cladistic analysis of the higher systematic categories of the Diapsida. Thesis (PhD in Palaeontology), University of California at Berkeley.
- Gauthier, J. A. 1986. Saurischian Monophyly and the Origin of Birds. In Padian, K. (Ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8: 1–55.
- Gauthier, J. & Padian, K. 1985. Phylogenetic, functional, and aerodynamic analyses of the origin of birds and their flight. In M. K. Hecht, J. H. Ostrom, G. Viohl, & P. Wellnhofer (eds.), The Beginnings of Birds pp. 185-197. Freunde des Jura-Museums, Eichstaett.
- Holtz, T. R., Jr., Molnar, R. E. & Currie, P. J. 2004. Basal Tetanurae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (eds.), The Dinosauria Second Edition. University of California Press, Berkeley: 71-110.
- Holtz, T. R., Jr., & Osmólska, H. 2004. Saurischia. In Weishampel, D. B., Dodson, P. & Osmólska, H. (eds.), The Dinosauria Second Edition. University of California Press, Berkeley: 21-24.
- Hunt, A. & Lucas, S. 1987. J. W. Stovall and the Mesozoic of the Cimmarron Valley, Oklahoma and New Mexico. New Mexico Geological Society Guidebook, 38th Field Conference, Northeastern New Mexico: 139-151.
- Janensch, W. 1950b. Die Skelettrekonstruktion von Brachiosaurus brancai. Palaeontographica (Suppl. 7) 3: 97-103 (in German).
- Kurtén, B. 1968. The Age of Dinosaurs. Weidenfeld & Nicolson, London.
- Kurzanov, S. M. 1987. Avimimidae and the problem of the origin of birds. Transactions of the Joint Soviet-Mongolian Paleontological Expedition 31:5-92.
- Langer, M. C. 2004. Basal Saurischia. In D. B. Weishampel, P. Dodson & H. Osmólska (eds.), The Dinosauria Second Edition. University of California Press, Berkeley: 25-46.
- Langer, M. C., Ezcurra, M. D., Bittencourt, J. S. & Novas, F. E. 2009. The origin and early evolution of dinosaurs. Biological Reviews 84:1-56.
- Longrich, N. 2006. Structure and function of hindlimb feathers in Archaeopteryx lithographica. Paleobiology 32 (3):417-431.
- Lucas, S. G., Herne, M. C., Heckert, A. B., Hunt, A. G. & Sullivan, R. M. 2004. Reappraisal of Seismosaurus, a Late Jurassic sauropod dinosaur from New Mexico. Geological Society of America Abstracts with Programs 36: 422.
- Luo, Z.-X., Kielan-Jaworowska, Z. & Cifelli, R. L. 2002. In quest for a phylogeny of Mesozoic mammals. Acta Palaeontologica Polonica 47: 1-78.
- Makovicky, P.J., Kobayashi, Y. & Currie, P.J. 2004. Ornithomimosauria. In D. B. Weishampel, P. Dodson & H. Osmólska (eds.), The Dinosauria, Second Edition. University of California Press, Berkeley: 137-150.
- Martinez, R. N. & Alcober, O. A. 2009. A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS ONE 4:1–12.
- Molnar, R. E., Kurzanov, S. M. & Dong, Z. 1990. Carnosauria. In D. B. Weishampel, Dodson & H. Osmólska (eds.), The Dinosauria. University of California Press, Berkeley: 169-209.
- Norman, D. B., Witmer, L. M. & Weishampel, D. B. 2004. Basal Ornithischia. In D. B. Weishampel, P. Dodson & H. Osmólska (eds.), The Dinosauria Second Edition, University of California Press, Berkeley: 325-334.
- Novas, F. E. 1992a. Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae. Palaeontology 35:51-62.
- Novas, F. E. 1994. New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina. Journal of Vertebrate Paleontology 13:400-423.
- Osmólska, H. & Roniewicz, E. 1970. Deinocheiridae, a new family of theropod dinosaurs. Palaeontologica Polonica 21:5-19.
- Ostrom, J. H. 1972. Dinosaur. In McGraw-Hill Yearbook of Science and Technology. McGraw-Hill, New York: 176-179.
- Ostrom, J. H. 1987. Protoavis, a Triassic bird? Archaeopteryx 5: 113-114.
- Ostrom, J. H. 1991. The bird in the bush. Nature 353 (6341):212.
- Padian, K. 1982. Macroevolution and the origin of major adaptations: Vertebrate flight as a paradigm for the analysis of patterns. In B. L. Mamet & M. J. Copeland (eds.), Third North American Palaeontological Convention Proceedings. North American Paleontological Convention, Montreal. Vol.2: 387-392.
- Padian, K. & May, C.L. 1993. The earliest dinosaurs. In S. G. Lucas & M. Morales (eds.), The nonmarine Triassic, New Mexico Museum of Natural History and Science Bulletin 3:379-382.
- Patterson, C. 1993. Naming names. Nature 366:518.
- Paul, G.S. 1984a. The segnosaurian dinosaurs: Relics of the prosauropod-ornithischian transition? Journal of Vertebrate Paleontology 4:507-515.
- Paul, G. S. 1988a. Predatory Dinosaurs of the World: A Complete Illustrated Guide. Simon and Schuster, New York, 464 p.
- Paul, G. S. 1988b. The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world’s largest dinosaurs. Hunteria 2 (3): 1-14.
- Paul, G. S. 2002. Dinosaurs of the Air: the Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press, Baltimore and London, i-ix + 460 p.
- Pettigrew, J. D. 1986. Flying primates? Megabats have the advanced pathway from eyes to midbrain. Science 231:1304-1306.
- Pettigrew, J. D., Jamieson, B. G. M., Robson, S. K., Hall, L. S., McAnally, K. I. & Cooper, H. M. Phylogenetic relations between microbats, megabats and primates (Mammalia: Chiroptera and Primates). Philosophical Transactions of the Royal Society of London B 325:489-559.
- Pisani, D., Yates, A. M., Langer, M. & Benton, M. J. 2002. A genus-level supertree of the Dinosauria. Proceedings of the Royal Society B 269:915-921.
- Prum, R. O. 1999. Development and Evolutionary Origin of Feathers. Journal of Experimental Zoology (Molecular and Developmental Evolution) 285: 291-306.
- Prum, R. O. & Brush, A. H. 2002. The evolutionary origin and diversification of feathers. Quarterly Review of Biology. 77: 261-295.
- Rauhut, O. W. M. 1995. The systematic position of the African theropods Carcharodontosaurus Stromer 1931 and Bahariasaurus Stromer 1934. Berliner Geowissenschaftliche Abhandlungen, E, 16.1:357-375 (in German).
- Rauhut, O. W. M., Milner, A. C. & Moore-Fay, S. 2009. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zoological Journal of the Linnean Society.
- Ray, G. E. 1941. Big for his day. Natural History 48:36-39.
- Reed, C.A. 1960. Polyphyletic or monophyletic ancestry of mammals, or: what is a class? Evolution 14: 314–322.
- Salgado, L., Coria, R. A. & Calvo, J. O. 1997. Evolution of titanosaurid sauropods. Part I. Phylogenetic analysis based on the postcranial evidence. Ameghiniana 34:3-32.
- Senter, P. 2006. Scapular orientation in theropods and basal birds, and the origin of flapping flight. Acta Palaeontologica Polonica 51 (2): 305–313.
- Sereno, P.C. & Arcucci, A.B. 1994. Dinosaur precursors from the Middle Triassic of Argentina: Lagerpeton chanarensis. Journal of Vertebrate Paleontology 13:385-399.
- Sereno, P. C., Rao, C. & Li, J. 2002. Sinornis satensis (Aves: Enantiornithes), from the Early Cretaceous of Northeastern China. In: Chiappe, L. M. & Witmer, L. M. (eds.), Mesozoic Birds, Above the Heads of Dinosaurs, University of California Press: 184-208.
- Simmons, N. B. 1994. The case for chiropteran monophyly. American Museum Novitates 3103: 54 pp.
- Simpson, G. G. 1959. Mesozoic mammals and the polyphyletic origin of mammals. Evolution 13: 405–414.
- Smith, D. K. 1996. A discriminant analysis of Allosaurus population using quarries as the operational units. Museum of Northern Arizona Bulletin 60:69-72.
- Smith, D. K. 1999. Patterns of size-related variation within Allosaurus. Journal of Vertebrate Paleontology 19 (2):402-403.
- Smith, N. D. & Pol, D. 2007. Anatomy of a basal sauropodomorph dinosaur from the Early Jurassic Hanson Formation of Antarctica. Acta Palaeontologica Polonica 52 (4): 657–674.
- Taylor, M. P. 2009. A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914). Journal of Vertebrate Paleontology 29 (3):787-806.
- Taylor, M. P., Wedel, M. J. & Naish, D. 2009. Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica 54(2): 213-220.
- Thulborn, R. A. 1975. Dinosaur polyphyly and the classification of archosaurs and birds. Australian Journal of Zoology 23:249-270.
- Thulborn, R. A. 1984. The avian relationships of Archaeopteryx, and the origin of birds. Zoological Journal of the Linnean Society 82:119-158.
- Upchurch, P. 1995. Evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London B 349: 365–390.
- Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124: 43–103.
- Upchurch, P., Barrett, P. M., Zhao, X.-J. & Xu, X. 2007b. A re-evaluation of Chinshakiangosaurus chunghoensis Ye vide Dong 1992 (Dinosauria, Sauropodomorpha): implications for cranial evolution in basal sauropod dinosaurs. Geological Magazine 144: 247–262.
- Wellnhofer, P. 1992. Protoavis - The oldest bird? Naturwissenschaftliche Rundschau 45(3):107-108 (in German).
- Wilson, J. A. & Sereno, P. C. 1998. Early evolution and higher level phylogeny of sauropod dinosaurs. Society of Vertebrate Paleontology Memoir 5: 1–68; Suppl. to Journal of Vertebrate Paleontology 18 (2).
- Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136:217–276.
- Xu, X. & Guo, Y. 2009. The origin and early evolution of feathers: insights from recent paleontological and neontological data. Vertebrata PalAsiatica 47 (4): 311-329.
- Xu, X., Zhao, X. & Clark, J. M. 2001. A new therizinosaur from the Lower Jurassic Lower Lufeng Formation of Yunnan, China. Journal of Vertebrate Paleontology 21 (3):477-483.
- Mareš, J. 1993. Záhada dinosaurů. Nakladatelství Svoboda-Libertas, Praha, 286 p.
- Sarvaš, R. 2003. Lidi zachvátila dinosauří horečka. 21. Století 8/2003, 82-85.



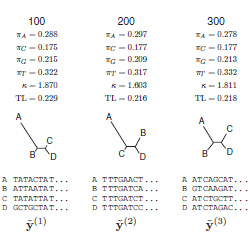




.png)



Parádní a netypicky pojatý článek hned na úvod - donutils mě Záhadu znovu vytáhnout, i když jsem ji četl po x-té někdy v září ;o). Musím říct, že v drtivé většině bodů se v názoru shodneme, vadí mi jak neopodstatněné nadsazování rozměrů (protože tady je to v několika případech čistě svéhlavá Marešova akce - a tos ještě nezmínil 17-20 metrové saurolofy a tarbosaury ;)). Ruší mě upřímně řečeno i ta závěrečná krypto-pasáž, dal bych tam možná jeden odstavec na tohle téma, ale prostě po přečtení celé knihy mi to tam nepasuje - chápu ale, že je to věc gusta a Marešovi to těžko s jeho zaměřením mít za zlé. Hlavním problémem (z hlediska faktografického)je asi to - a z tvého článku to plyne zcela zřetelně - dost úzký souhrn zdrojů, se kterým Mareš pracoval - v podstatě Paul, Bakker, sem tam nějaký Gauthier, odkaz na osobní komentář Currieho - a tím to vlastně hasne; na podobně obecně pojatou knihu to asi přece jen chtělo větší penzum zdrojů - nevěřím tomu, že by si je neobstaral, vždyť kontakty na to měl. Pak by asi spousta věcí vypadala jinak, nad tím už ale nemá smysl spekulovat. I já ale Záhadu beru veskrze kladně a dlouho to byl jeden z mých hlavních informačních zdrojů, nad čímž se dneska člověk už musí jen pousmát... Spíš je k zamyšlení, že ačkoli ZD měla daleko od odborně paleontologické knihy, byla v našich končinách dlouho v podstatě tím nejlepším, co na téma dinosaurů vyšlo - to je skutečně politováníhodné a svědčí to o tom, jak jsme byli pozadu za zahraničím (dnes se to naštěstí všechno změnilo s příchodem Internetu). To ale nemění nic na tom, že mám Záhadu stále na čestném místě na poličce (už jen, kdyby nic jiného, než že Mareš zmínil poslední závěry Gauthiera z r. 86 a v samotném závěru i Mary Schwetzer, což byla skutečně tehdy informace maximálně aktuální, má u mě palec nahoru). Jinak ale skvělý článek, Davide ;o)
OdpovědětVymazatJiří M.
P.S. Ten psittacoceratops v pasáži o savcích má předpkládám být psittacosaurus ;o)
Ou...to se tam dostalo? Už je to každopádně opravené. :o) Jinak ano, souhlasím. Bakker a Paul byli často předvídaví až neuvěřitelně (což nemění nic na tom, že v jejich době byla pro to, co psali, často nedostatečná podpora), ale ve spoustě jiných záležitostí, ve kterých nesouhlasili se závěry své doby, pro změnu pravdu neměli. V poslední době musím třeba vyloženě žasnout, jak to Paulovi vychází - evidence pro to, že deinonychosauři jsou druhotně nelétaví je teď velmi silná (viz třeba citovaný Longrich 2006), stejně jako pro to, že plachtění u čtyřkřídlých forem je skutečně to, co předcházelo aktivnímu letu.
OdpovědětVymazatTaky si myslím, že silná pozice Záhady ve zdejších končinách je už dnes trochu bezdůvodná - konec konců to píšu v článku, ta kniha sice krásně konzervuje názory určitých autorů v určité době, ale dnes už je neaktuální. Dan na staré verzi blogu psal, že je to doporučená kniha i ve vysokoškolských skriptech, a to skutečně není v pořádku.
BTW, pokud by se nějaký článek skutečně líbil, můžeš využít i možnosti hodnocení hvězdičkami dole (jedna z novinek); tomu ale nepřikládám velkou důležitost. Každopádně díky za pochvalné hodnocení, budu se snažit úroveň udržet. ;o)